Potrebbero piacerti anche
- Tumor Suppressor APC Blocks DNA Polymerase - Dependent Strand Displacement Synthesis During Long Patch But Not Short Patch Base Excision Repair and Increases Sensitivity To Methylmethane SulfonateDocumento8 pagineTumor Suppressor APC Blocks DNA Polymerase - Dependent Strand Displacement Synthesis During Long Patch But Not Short Patch Base Excision Repair and Increases Sensitivity To Methylmethane SulfonateBiosynthesisNessuna valutazione finora
- And + Peptide That Is Recognized by Human CD4 Characterization of ADocumento13 pagineAnd + Peptide That Is Recognized by Human CD4 Characterization of ABiosynthesisNessuna valutazione finora
- Induction of Hepatitis A Virus-Neutralizing Antibody by A Virus-Specific Synthetic PeptideDocumento4 pagineInduction of Hepatitis A Virus-Neutralizing Antibody by A Virus-Specific Synthetic PeptideBiosynthesisNessuna valutazione finora
- A Simple Method For Displaying The Hydropathic Character of A ProteinDocumento28 pagineA Simple Method For Displaying The Hydropathic Character of A ProteinBiosynthesisNessuna valutazione finora
- Silencing of microRNAs in Vivo With Antagomirs'Documento5 pagineSilencing of microRNAs in Vivo With Antagomirs'BiosynthesisNessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
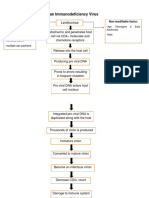
- Human Immunodeficiency Virus: LentiruvirusDocumento5 pagineHuman Immunodeficiency Virus: Lentiruvirusjoyrena ochondraNessuna valutazione finora
- 2010 11 18 - Sex On The HillDocumento13 pagine2010 11 18 - Sex On The HillThe University Daily Kansan100% (2)
- Liver Biopsy - H. Takahashi (Intech, 2011) WWDocumento414 pagineLiver Biopsy - H. Takahashi (Intech, 2011) WWmientweg100% (1)
- Polemonium Reptans Euterpe Oleracea: Aloe VeraDocumento11 paginePolemonium Reptans Euterpe Oleracea: Aloe VeraSarah McOnellyNessuna valutazione finora
- VBI Vaccines - ICAAC Presentation 2014Documento12 pagineVBI Vaccines - ICAAC Presentation 2014DougNessuna valutazione finora
- Proteus Mirabilis Infections - StatPearls - NCBI BookshelfDocumento5 pagineProteus Mirabilis Infections - StatPearls - NCBI BookshelfclaudyNessuna valutazione finora
- Chapter 8 NematodesDocumento5 pagineChapter 8 NematodesJuju GalangNessuna valutazione finora
- Animal Diseases PDFDocumento8 pagineAnimal Diseases PDFSubash KrishnanNessuna valutazione finora
- Drug StudyDocumento11 pagineDrug StudyKimberly Ann MendozaNessuna valutazione finora
- Neonatal Infections Part 1 - MDM DygDocumento30 pagineNeonatal Infections Part 1 - MDM DygNana YunusNessuna valutazione finora
- 16 MLT PDFDocumento148 pagine16 MLT PDFTanveerNessuna valutazione finora
- Micro-Para Lesson 1Documento11 pagineMicro-Para Lesson 1Shene Claire VigillaNessuna valutazione finora
- 35-1404729796 JurnalDocumento3 pagine35-1404729796 JurnalNancy ZhangNessuna valutazione finora
- Para Lec CombinedDocumento83 paginePara Lec CombinedClent Earl Jason O. BascoNessuna valutazione finora
- Doctor Interviews Documentaries & Research QuotesDocumento32 pagineDoctor Interviews Documentaries & Research QuotesjdhNessuna valutazione finora
- Nose, Sinuses, Mouth, and LarynxDocumento3 pagineNose, Sinuses, Mouth, and LarynxjottowagNessuna valutazione finora
- Bacterial Disease Mechanisms: An Introduction To Cellular MicrobiologyDocumento685 pagineBacterial Disease Mechanisms: An Introduction To Cellular MicrobiologyVero VillarrealNessuna valutazione finora
- Cattle Diseases Farmers GuideDocumento77 pagineCattle Diseases Farmers GuideDivyajyoti DevaNessuna valutazione finora
- Chapter 22 Upper Respiratory DisorderDocumento171 pagineChapter 22 Upper Respiratory DisorderAnnette Aquino GuevarraNessuna valutazione finora
- Viral Infections of The Gastrointestinal Tract and Viral Infections of The Genitourinary SystemDocumento14 pagineViral Infections of The Gastrointestinal Tract and Viral Infections of The Genitourinary SystemDARLENE CLAIRE ANDEZANessuna valutazione finora
- Art Integrated Project of Biology Class XII-BDocumento10 pagineArt Integrated Project of Biology Class XII-BRonit AdhikaryNessuna valutazione finora
- Malabsorption Syndrome in ChickensDocumento5 pagineMalabsorption Syndrome in ChickensŠhâh NawazNessuna valutazione finora
- ChromID CPS Brochure Final-1Documento2 pagineChromID CPS Brochure Final-1horia96Nessuna valutazione finora
- Review On Dermatomycosis Jurnal 2Documento23 pagineReview On Dermatomycosis Jurnal 2Laela NurrochmahNessuna valutazione finora
- MicrobiologyDocumento17 pagineMicrobiologykimacol659Nessuna valutazione finora
- Unit 2 - Innovations in Health and Medicine (Paragraphs)Documento1 paginaUnit 2 - Innovations in Health and Medicine (Paragraphs)April ThunNessuna valutazione finora
- Hydronephrosis LWDocumento8 pagineHydronephrosis LWvitauxianaNessuna valutazione finora
- Nalanda Vidyalaya - Colombo 10: 2 Term - Unit Test Project Project English LanguageDocumento2 pagineNalanda Vidyalaya - Colombo 10: 2 Term - Unit Test Project Project English Languagesaksi76@gmail.comNessuna valutazione finora
- BasicMicrobiology DrVillavisencio Aug2019Documento66 pagineBasicMicrobiology DrVillavisencio Aug2019Mi ColesNessuna valutazione finora
- ChikungunyaDocumento2 pagineChikungunyaDicky Budi NurcahyaNessuna valutazione finora