Potrebbero piacerti anche
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- 11th House of IncomeDocumento9 pagine11th House of IncomePrashanth Rai0% (1)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Understanding Death Through the Life SpanDocumento52 pagineUnderstanding Death Through the Life SpanKaren HernandezNessuna valutazione finora
- Philippine Bank of Communications Vs Commissioner of Internal RevenueDocumento2 paginePhilippine Bank of Communications Vs Commissioner of Internal RevenueNFNL100% (1)
- Margiela Brandzine Mod 01Documento37 pagineMargiela Brandzine Mod 01Charlie PrattNessuna valutazione finora
- Brother LS2300 Sewing Machine Instruction ManualDocumento96 pagineBrother LS2300 Sewing Machine Instruction ManualiliiexpugnansNessuna valutazione finora
- Joey Cena 2Documento1 paginaJoey Cena 2api-635313033Nessuna valutazione finora
- Manoj KR - KakatiDocumento5 pagineManoj KR - Kakatimanoj kakatiNessuna valutazione finora
- Hold-Up?" As He Simultaneously Grabbed The Firearm of Verzosa. WhenDocumento2 pagineHold-Up?" As He Simultaneously Grabbed The Firearm of Verzosa. WhenVener MargalloNessuna valutazione finora
- NGOs Affiliated To SWCDocumento2.555 pagineNGOs Affiliated To SWCMandip marasiniNessuna valutazione finora
- David Emmett & Graeme Nice - What You Need To Know About Cannabis - Understanding The FactsDocumento120 pagineDavid Emmett & Graeme Nice - What You Need To Know About Cannabis - Understanding The FactsJovana StojkovićNessuna valutazione finora
- Nursing ManagementDocumento14 pagineNursing ManagementNolan Ivan EudinNessuna valutazione finora
- B§ÐmMm OÝ_ : EH$ {Z~§YmVrc H$ënZmDocumento70 pagineB§ÐmMm OÝ_ : EH$ {Z~§YmVrc H$ënZmVikas NikharangeNessuna valutazione finora
- Focus2 2E Review Test 4 Units1 8 Vocabulary Grammar UoE Reading GroupBDocumento4 pagineFocus2 2E Review Test 4 Units1 8 Vocabulary Grammar UoE Reading GroupBaides1sonNessuna valutazione finora
- Choosing the Right Organizational Pattern for Your SpeechDocumento19 pagineChoosing the Right Organizational Pattern for Your SpeechKyle RicardoNessuna valutazione finora
- Mock-B1 Writing ReadingDocumento6 pagineMock-B1 Writing ReadingAnonymous 0uBSrduoNessuna valutazione finora
- 41720105Documento4 pagine41720105renu tomarNessuna valutazione finora
- Facebook Use Case Diagram Activity Diagram Sequence DiagramDocumento21 pagineFacebook Use Case Diagram Activity Diagram Sequence DiagramSaiNessuna valutazione finora
- Eng CBLDocumento2 pagineEng CBLMengTangNessuna valutazione finora
- Tarea 1Documento36 pagineTarea 1LUIS RVNessuna valutazione finora
- MA KP3-V2H-2 enDocumento155 pagineMA KP3-V2H-2 enJavier MiramontesNessuna valutazione finora
- HIstory of PerfumeDocumento3 pagineHIstory of PerfumebetselevenNessuna valutazione finora
- Biological Control in Brazil An OverviewDocumento10 pagineBiological Control in Brazil An OverviewGustavo Ferreira MoraisNessuna valutazione finora
- Semester 3 SyllabusDocumento12 pagineSemester 3 SyllabusFuggi JaanNessuna valutazione finora
- Commissioning Procedure for JTB-PEPCDocumento17 pagineCommissioning Procedure for JTB-PEPCelif maghfirohNessuna valutazione finora
- 6 Economics of International TradeDocumento29 pagine6 Economics of International TradeSenthil Kumar KNessuna valutazione finora
- Kimone Wright - Registered Nurse ResumeDocumento2 pagineKimone Wright - Registered Nurse Resumeapi-365123958Nessuna valutazione finora
- Colorectal Disease - 2023 - Freund - Can Preoperative CT MR Enterography Preclude The Development of Crohn S Disease LikeDocumento10 pagineColorectal Disease - 2023 - Freund - Can Preoperative CT MR Enterography Preclude The Development of Crohn S Disease Likedavidmarkovic032Nessuna valutazione finora
- Ce QuizDocumento2 pagineCe QuizCidro Jake TyronNessuna valutazione finora
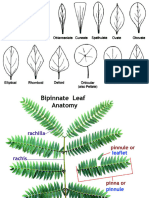
- Types, Shapes and MarginsDocumento10 pagineTypes, Shapes and MarginsAkhil KanukulaNessuna valutazione finora
- Week 10 8th Grade Colonial America The Southern Colonies Unit 2Documento4 pagineWeek 10 8th Grade Colonial America The Southern Colonies Unit 2santi marcucciNessuna valutazione finora