Potrebbero piacerti anche
- Cuadro de Vias MetabolicasDocumento2 pagineCuadro de Vias MetabolicasAmairany Escudero Cifuentes85% (13)
- Estabilidad de EmulsionesDocumento0 pagineEstabilidad de EmulsionesVictor HernandezNessuna valutazione finora
- Rutas Metabolicas - BCTDocumento4 pagineRutas Metabolicas - BCTLizSR01100% (3)
- Infografía Lipogénesis Vs LipólisisDocumento14 pagineInfografía Lipogénesis Vs Lipólisismariana opolka100% (2)
- Carbohidratos ConejoDocumento5 pagineCarbohidratos Conejomonica100% (1)
- Enfermedades Causadas Por Afectaciones en Vías MetabólicasDocumento6 pagineEnfermedades Causadas Por Afectaciones en Vías MetabólicasMarianaNessuna valutazione finora
- GLUCÓLISISDocumento18 pagineGLUCÓLISISOscar Antunez BitesNessuna valutazione finora
- Balanza Heuristica G10-ComprimidoDocumento19 pagineBalanza Heuristica G10-ComprimidoLAURA HAYDEE CASTILLO DONISNessuna valutazione finora
- Fly RockDocumento19 pagineFly Rockllllcesarllll100% (1)
- Bioquímica Digestión de CarbohidratosDocumento4 pagineBioquímica Digestión de CarbohidratosRafael ColmenaresNessuna valutazione finora
- Ciclo de KrebsDocumento6 pagineCiclo de KrebsAguila PaulNessuna valutazione finora
- 5.2 Tabla de Vías MetabólicasDocumento3 pagine5.2 Tabla de Vías MetabólicasRebeca RioNessuna valutazione finora
- Demostracion de Cuerpos Cetonicos en La OrinaDocumento21 pagineDemostracion de Cuerpos Cetonicos en La OrinaYiselda LisetheNessuna valutazione finora
- Biosíntesis de ColesterolDocumento6 pagineBiosíntesis de ColesterolAlejandra ViteriNessuna valutazione finora
- Tabla de Las Rutas MetabólicasDocumento4 pagineTabla de Las Rutas MetabólicasJavier Vázquez OrdoñezNessuna valutazione finora
- Metabolismo Del GlucogenoDocumento34 pagineMetabolismo Del GlucogenoEsmeralda CarreraNessuna valutazione finora
- 1.metabolismo Específico de Los Tejidos 2016Documento62 pagine1.metabolismo Específico de Los Tejidos 2016Junior Uriarte DávilaNessuna valutazione finora
- Cetolisis CetogenesisDocumento10 pagineCetolisis CetogenesisTania Huarcaya100% (1)
- LIPOLISISDocumento8 pagineLIPOLISISScarlette OlivoNessuna valutazione finora
- Metabolismo de Hemoglobina (DV)Documento9 pagineMetabolismo de Hemoglobina (DV)Dary Deivy Varela AracheNessuna valutazione finora
- GLUCOGENOLISISDocumento6 pagineGLUCOGENOLISISCristobal Gamboa CamposNessuna valutazione finora
- GluconeogénesisDocumento16 pagineGluconeogénesisGiovanni GonzalezNessuna valutazione finora
- Funciones No Respiratorias Del PulmónDocumento23 pagineFunciones No Respiratorias Del PulmónCata EnlacesNessuna valutazione finora
- Sintesis de TrigliceridosDocumento5 pagineSintesis de Trigliceridoskatiyerar100% (3)
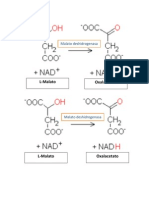
- La Enzima Malato DeshidrogenasaDocumento2 pagineLa Enzima Malato DeshidrogenasaKaren Torres JuárezNessuna valutazione finora
- Balanza Heuristica Comparacion Glucolisis y GluconeogenesisDocumento4 pagineBalanza Heuristica Comparacion Glucolisis y GluconeogenesisESTRELLA CELESTE AVENDA�O DIAZNessuna valutazione finora
- Sintesis, Transporte y Excrecion de ColesterolDocumento4 pagineSintesis, Transporte y Excrecion de ColesterolPao Condo HidalgoNessuna valutazione finora
- Metabolismo Del EritrocitoDocumento11 pagineMetabolismo Del EritrocitoJGQM08Nessuna valutazione finora
- Gluconeogenesis y Control de La Glucosa en SangreDocumento37 pagineGluconeogenesis y Control de La Glucosa en SangreStefano GarciaNessuna valutazione finora
- Fase No Oxidativa Pentosas Fosfatos CompletaDocumento21 pagineFase No Oxidativa Pentosas Fosfatos Completaartu0oo0% (1)
- Mi Lipogenesis 2018 Final PDFDocumento34 pagineMi Lipogenesis 2018 Final PDFAna HurtadoNessuna valutazione finora
- Actividad en Clase de Cuerpos CetonicosDocumento5 pagineActividad en Clase de Cuerpos CetonicosEzequiel SalazarNessuna valutazione finora
- Oxidacion de Ácidos GrasosDocumento1 paginaOxidacion de Ácidos Grasosagustin zarate cruzNessuna valutazione finora
- Acetil CoenzimaDocumento3 pagineAcetil CoenzimaJose Dominguez100% (1)
- Esofago y EstomagoDocumento6 pagineEsofago y EstomagoCarlos ValenzuelaNessuna valutazione finora
- CetogenesisDocumento8 pagineCetogenesisYuukiAuclair100% (1)
- Regulación de La GlucólisisDocumento3 pagineRegulación de La Glucólisisfrancisco lozano gomezNessuna valutazione finora
- Cuadro ComparativoDocumento3 pagineCuadro ComparativoDenisse PerezNessuna valutazione finora
- LIPOPROTEINASDocumento15 pagineLIPOPROTEINASJhon Eswin Lopez BernalNessuna valutazione finora
- Glosario de Terminos PsiquiátricosDocumento2 pagineGlosario de Terminos PsiquiátricosJeani LascanoNessuna valutazione finora
- Metabolismo de CarbohidratosDocumento11 pagineMetabolismo de CarbohidratosJuan Ernesto AnguloNessuna valutazione finora
- Lanzaderas MitocondrialesDocumento3 pagineLanzaderas MitocondrialesJoseNeiserCastilloArrascue0% (1)
- Exposiciones Gluconeogenesis 11Documento22 pagineExposiciones Gluconeogenesis 11Hernan Mauricio Rivera Escobar50% (2)
- La GlucogenesisDocumento4 pagineLa GlucogenesisJonathan Paul Otoya BarturenNessuna valutazione finora
- Gluconeogénesis - Ciclo de Las PentosasDocumento57 pagineGluconeogénesis - Ciclo de Las PentosasUSMP FN ARCHIVOSNessuna valutazione finora
- Metabolismo Del GlucogenoDocumento30 pagineMetabolismo Del GlucogenoMAURICIO AUGUSTO ORELLANA CARDOZO100% (1)
- Ciclo Del Ácido TricarboxílicoDocumento32 pagineCiclo Del Ácido TricarboxílicoDenissLopez100% (1)
- Ciclo de CoriDocumento7 pagineCiclo de CoriJeanette PaulinaNessuna valutazione finora
- Glucolisis 21645 PDFDocumento77 pagineGlucolisis 21645 PDFShärlÿ ContrerasNessuna valutazione finora
- LIPOPROTEINASDocumento6 pagineLIPOPROTEINASRuben Martinez100% (2)
- AGAR LIA 9.docxNNDocumento2 pagineAGAR LIA 9.docxNNAndrea CohenNessuna valutazione finora
- Hipertrigliceridemia EndógenaDocumento10 pagineHipertrigliceridemia EndógenaJosé LuisNessuna valutazione finora
- GluconeogenesisDocumento20 pagineGluconeogenesisdan100% (2)
- Enfermedad Von GierkeDocumento5 pagineEnfermedad Von GierkeMerceLópezNessuna valutazione finora
- Mapa Conceptual Metabolismo de Los Carbohidratos. Monserrat HernandezDocumento1 paginaMapa Conceptual Metabolismo de Los Carbohidratos. Monserrat Hernandezhernandezmonse684Nessuna valutazione finora
- Echinochloa PolystachyaDocumento9 pagineEchinochloa Polystachyaanon_993623997Nessuna valutazione finora
- Anatomía Pulmonar - 19 Abril 2023 - Dr. ChávezDocumento6 pagineAnatomía Pulmonar - 19 Abril 2023 - Dr. Chávezalguiensoy9512Nessuna valutazione finora
- Exposicion LipolisisDocumento9 pagineExposicion LipolisisMicrored Hospital LamasNessuna valutazione finora
- Apuntes Del Ciclo de KrebsDocumento13 pagineApuntes Del Ciclo de KrebsHerbertNessuna valutazione finora
- Vías MetabólicasDocumento24 pagineVías MetabólicasGabriela ZevallosNessuna valutazione finora
- Evidencia GlicolisisDocumento7 pagineEvidencia GlicolisisArely LopezNessuna valutazione finora
- Teorico 4 Metabolismo de GlucidosDocumento25 pagineTeorico 4 Metabolismo de GlucidosAdriano pompeiNessuna valutazione finora
- Capitulo 2 (Astronomia)Documento84 pagineCapitulo 2 (Astronomia)Yackelyn Pastor100% (1)
- Proy Final - JorleniDocumento29 pagineProy Final - JorleniEduardo Salazar RNessuna valutazione finora
- Dos ComponentesDocumento26 pagineDos ComponentesNelida AgramontNessuna valutazione finora
- Topografia GeneralDocumento29 pagineTopografia GeneralDaniel HanoverNessuna valutazione finora
- Función AfinDocumento3 pagineFunción AfinCesar GomezNessuna valutazione finora
- Informe 1 Colorantes Naturales PDFDocumento25 pagineInforme 1 Colorantes Naturales PDFErick LopezNessuna valutazione finora
- Cizallado PDFDocumento10 pagineCizallado PDFleogrignafini100% (1)
- Método de La Regla FalsaDocumento2 pagineMétodo de La Regla FalsaJavier Salvador Martinez SalasNessuna valutazione finora
- Tarea 1Documento4 pagineTarea 1DavidNessuna valutazione finora
- CORRIGIENDO Estudio Mecanica de Suelos ZDocumento61 pagineCORRIGIENDO Estudio Mecanica de Suelos ZZelaya WilsonNessuna valutazione finora
- Inversor PDFDocumento32 pagineInversor PDFEverDorantesANessuna valutazione finora
- Interpolación Polinomial de Newton en Diferencias DivididasDocumento7 pagineInterpolación Polinomial de Newton en Diferencias DivididasBladimir Perez SilvaNessuna valutazione finora
- El Proceso de MedDocumento55 pagineEl Proceso de MedAlejandro HernandezNessuna valutazione finora
- Consideraciones en La Aplicación Del MTMDocumento4 pagineConsideraciones en La Aplicación Del MTMbaro4518Nessuna valutazione finora
- Imaginarios AtávicosDocumento4 pagineImaginarios AtávicosMaria PerezNessuna valutazione finora
- Diseño ExperimentalDocumento17 pagineDiseño ExperimentalgimenNessuna valutazione finora
- TanquesDocumento4 pagineTanquesAngel Stiven Romero DiazNessuna valutazione finora
- DISEÑO+DEDocumento15 pagineDISEÑO+DEWilliam García SánchezNessuna valutazione finora
- Control 3 Límites y DerivadasDocumento2 pagineControl 3 Límites y DerivadasAxlBassiNessuna valutazione finora
- Diseño y Construcción de Un Exoesqueleto Ligero Extensor de Fuerza 0Documento10 pagineDiseño y Construcción de Un Exoesqueleto Ligero Extensor de Fuerza 0polNessuna valutazione finora
- Volumen Gaseoso MolarDocumento12 pagineVolumen Gaseoso MolarCaroldo CaballeroNessuna valutazione finora
- Cap 10 Secc 10 6 Ecuaciones Polares de Las ConicasDocumento32 pagineCap 10 Secc 10 6 Ecuaciones Polares de Las Conicasyucra carbajal rubenNessuna valutazione finora
- Memoria DescriptivaDocumento4 pagineMemoria DescriptivajuliojcqfNessuna valutazione finora
- Transporte de SedimentosDocumento63 pagineTransporte de SedimentosKarencita Aycaya ChiqueNessuna valutazione finora
- Informe N 1 Uso Del Osciloscopio y Generador de Senales PDFDocumento16 pagineInforme N 1 Uso Del Osciloscopio y Generador de Senales PDFBryan ChallcoNessuna valutazione finora
- Informe Nanohub (Bastidas - Torres) PDFDocumento17 pagineInforme Nanohub (Bastidas - Torres) PDFmelito09Nessuna valutazione finora
- Edificio Prefabricado PDFDocumento98 pagineEdificio Prefabricado PDFjesus40Nessuna valutazione finora