Potrebbero piacerti anche
- Miología: sistema musculoesquelético: Guía de prácticasDa EverandMiología: sistema musculoesquelético: Guía de prácticasNessuna valutazione finora
- Adiestramiento Del PacienteDocumento26 pagineAdiestramiento Del PacienteKarina Herrera ZavaletaNessuna valutazione finora
- 2020 Texto KINE PDFDocumento120 pagine2020 Texto KINE PDFCristhian CoroNessuna valutazione finora
- Alteraciones Rotacionales en Miembros InferioresDocumento11 pagineAlteraciones Rotacionales en Miembros InferioresMayra Su100% (1)
- Herramientas de análisis para la estabilidad estática postural: Caso de aplicación: personas con amputación transtibialDa EverandHerramientas de análisis para la estabilidad estática postural: Caso de aplicación: personas con amputación transtibialNessuna valutazione finora
- Cambios Post-Morten en La CarneDocumento45 pagineCambios Post-Morten en La CarneJhurema B Ch67% (12)
- El nervio periférico: Estructura y funciónDa EverandEl nervio periférico: Estructura y funciónNessuna valutazione finora
- Estimulacion Electrica para Contraccion MuscularDocumento90 pagineEstimulacion Electrica para Contraccion MuscularPILAR HERRERANessuna valutazione finora
- GALVANISMODocumento5 pagineGALVANISMOyohalliNessuna valutazione finora
- Practica 4 Farmacologia Cardiovascular en Un Sistema SimuladoDocumento1 paginaPractica 4 Farmacologia Cardiovascular en Un Sistema Simuladojesus.checa.xoxotlaNessuna valutazione finora
- Articulo AmputadoDocumento10 pagineArticulo AmputadoLtf Oswaldo Guerrero IslasNessuna valutazione finora
- Sistema Muscular Embriologia 1Documento32 pagineSistema Muscular Embriologia 1jessyi1234550% (2)
- Osteoartrosis - Arevalo Fernandez Lizbeth AlexandraDocumento39 pagineOsteoartrosis - Arevalo Fernandez Lizbeth AlexandraTerapia Fisica Mejora JtlNessuna valutazione finora
- ANATOMIA Ligamento Cruzado AnteriorDocumento22 pagineANATOMIA Ligamento Cruzado Anteriorgeine LeliNessuna valutazione finora
- Miembro superior: Exposición, diagnóstico y prevención de desórdenes musculoesqueléticos en los ambientes laboralesDa EverandMiembro superior: Exposición, diagnóstico y prevención de desórdenes musculoesqueléticos en los ambientes laboralesNessuna valutazione finora
- Biomecanica en Fitness 1Documento80 pagineBiomecanica en Fitness 1Maik CaroNessuna valutazione finora
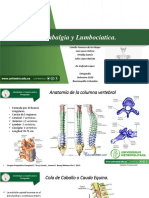
- Lumbalgia y LumbociaticaDocumento13 pagineLumbalgia y LumbociaticaCamilo Fonseca DlrNessuna valutazione finora
- Estructura y Funcion ArticularDocumento48 pagineEstructura y Funcion ArticularDimitriNessuna valutazione finora
- Método Pold® 2012Documento14 pagineMétodo Pold® 2012Juankrs Noom VargasNessuna valutazione finora
- Bloque Comun de Nivel II de Los Cursos de Entrenadores-As c-1 PDFDocumento347 pagineBloque Comun de Nivel II de Los Cursos de Entrenadores-As c-1 PDFXavi TeixidóNessuna valutazione finora
- Sistema Hematopoyético y Linfático TallerDocumento6 pagineSistema Hematopoyético y Linfático TallerSebastian ManriqueNessuna valutazione finora
- Lesiones Deportivas de HombroDocumento20 pagineLesiones Deportivas de HombroDANIEL ALEJANDRO100% (3)
- Polígono de WillisDocumento11 paginePolígono de WillisMariana Zavala BañuelosNessuna valutazione finora
- Fisiologia de La ContraccionDocumento41 pagineFisiologia de La ContraccionLizbeth Mp100% (3)
- 2014 Fracturas Maleolares Del Adulto y Luxaciones Del TobilloDocumento13 pagine2014 Fracturas Maleolares Del Adulto y Luxaciones Del TobilloFISIOKINESNessuna valutazione finora
- Tobillo PDFDocumento47 pagineTobillo PDFluchomena2702100% (3)
- Clase 5. Tejido MuscularDocumento43 pagineClase 5. Tejido MuscularAlejandro Martinez ManzanoNessuna valutazione finora
- Masokinesiterapia en Cirugia Estetica y ReconstructivaDocumento9 pagineMasokinesiterapia en Cirugia Estetica y Reconstructivajose ANessuna valutazione finora
- Anatomía y Fisiología Del Sistema ÓseoDocumento10 pagineAnatomía y Fisiología Del Sistema ÓseoPatty PerezNessuna valutazione finora
- Practico Generalidades 2021Documento11 paginePractico Generalidades 2021Alejandro MaureliaNessuna valutazione finora
- Métodos Específicos en Fisioterapia IIIDocumento10 pagineMétodos Específicos en Fisioterapia IIIjebf85Nessuna valutazione finora
- Enfermedades OseasDocumento65 pagineEnfermedades OseasAlejo Pérez Vinueza100% (1)
- Glosario de Fisioterapia Aula AbiertaDocumento11 pagineGlosario de Fisioterapia Aula AbiertaMiguel BorekNessuna valutazione finora
- Arteria SubclaviaDocumento37 pagineArteria SubclaviaCésar Martínez MorenoNessuna valutazione finora
- Consideraciones Tema EscoliosisDocumento5 pagineConsideraciones Tema Escoliosisporno331Nessuna valutazione finora
- Práctica de Tórax AnatomíaDocumento19 paginePráctica de Tórax AnatomíaPaulina Malinali Pérez BracamontesNessuna valutazione finora
- El Sistema Nervioso Es Muy ComplejoDocumento4 pagineEl Sistema Nervioso Es Muy Complejoalfmargon2737119Nessuna valutazione finora
- NeurotransmisoresDocumento17 pagineNeurotransmisoresReyna VaherNessuna valutazione finora
- LESION DEL MANGUITO ROTADOR CompletoDocumento52 pagineLESION DEL MANGUITO ROTADOR CompletoAngel LopezNessuna valutazione finora
- Programa de Salud EscoliosisDocumento11 paginePrograma de Salud EscoliosisJulia Elias SalamancaNessuna valutazione finora
- Generalidades de BiomecánicaDocumento5 pagineGeneralidades de BiomecánicaJessica AlexNessuna valutazione finora
- Biomecánica Del HuesoDocumento22 pagineBiomecánica Del HuesoAnthonyVelasquezMirandaNessuna valutazione finora
- Historia APTADocumento3 pagineHistoria APTAanama_170186% (7)
- Fisiologia Del EncefaloDocumento14 pagineFisiologia Del EncefaloAlexandra Amaya VanstrahlenNessuna valutazione finora
- Método de Intervención en La FisioterapiaDocumento13 pagineMétodo de Intervención en La FisioterapiaRebeca Jaimes MoncerattNessuna valutazione finora
- Antebrazo DistalDocumento16 pagineAntebrazo DistalEsteban CeruttiNessuna valutazione finora
- Proyecto de Fisioterapia!!!!!Documento15 pagineProyecto de Fisioterapia!!!!!willians diazNessuna valutazione finora
- Teoria de SistemasDocumento3 pagineTeoria de SistemasBetzabeth Gomez100% (2)
- Cerebelo AtaxiaDocumento45 pagineCerebelo AtaxianekotefiNessuna valutazione finora
- LumbociaticaDocumento11 pagineLumbociaticaTomas Bernardo Alarcon BenitezNessuna valutazione finora
- HidrokinesiterapiaDocumento29 pagineHidrokinesiterapiaINGRID JULIETH VILLAMIL TORRESNessuna valutazione finora
- Anatomía Descriptiva Sacro & CóccixDocumento38 pagineAnatomía Descriptiva Sacro & CóccixLauraPatriciaNessuna valutazione finora
- Tema 4. Fundamentos de FisioterapiaDocumento9 pagineTema 4. Fundamentos de FisioterapiaAna Diaz Vazquez100% (1)
- FNPDocumento7 pagineFNPIsrael PapinuttiNessuna valutazione finora
- Amputaciones Del Miembro SuperiorDocumento11 pagineAmputaciones Del Miembro SuperiorGonzalo Zegarra Ordoñez100% (2)
- Sistema MusculoesqueléticoDocumento8 pagineSistema MusculoesqueléticoMónica JiménezNessuna valutazione finora
- Psico Motricidad PreguntasDocumento15 paginePsico Motricidad PreguntashilmambNessuna valutazione finora
- Plexo CoccigeoDocumento1 paginaPlexo CoccigeoAylín Colina JordánNessuna valutazione finora
- FT ErgonomiaDocumento18 pagineFT ErgonomiaCopias DodiNessuna valutazione finora
- Red Mexicana de Municipios Por La SaludDocumento8 pagineRed Mexicana de Municipios Por La SaludFatima GonzálezNessuna valutazione finora
- Proyecto NoeDocumento25 pagineProyecto NoeGenesis GuachallaNessuna valutazione finora
- LumbarDocumento8 pagineLumbarKarina Herrera ZavaletaNessuna valutazione finora
- Amputación de Dedo Pulgar y ProtetizaciónDocumento18 pagineAmputación de Dedo Pulgar y Protetizaciónasddsa0% (1)
- Cuidado De Enfermería Corporal-Social a La Persona Posamputada Portadora De Dolor De Miembro Fantasma (Dmf)Da EverandCuidado De Enfermería Corporal-Social a La Persona Posamputada Portadora De Dolor De Miembro Fantasma (Dmf)Nessuna valutazione finora
- Power Anato 1Documento18 paginePower Anato 1DANIEL ALEJANDRONessuna valutazione finora
- Anatomia Aplicada 12º Sistema Nervioso Cerebro Vias 1Documento59 pagineAnatomia Aplicada 12º Sistema Nervioso Cerebro Vias 1DANIEL ALEJANDRO100% (1)
- Anatomia Aplicada 11º Sistema Nervioso Generalidades Parte 2Documento19 pagineAnatomia Aplicada 11º Sistema Nervioso Generalidades Parte 2DANIEL ALEJANDRONessuna valutazione finora
- Anatomia Aplicada 11º Sistema Nervioso Generalidades Parte 2Documento19 pagineAnatomia Aplicada 11º Sistema Nervioso Generalidades Parte 2DANIEL ALEJANDRONessuna valutazione finora
- Power Biomecanica N°3 LumbociaticasDocumento19 paginePower Biomecanica N°3 LumbociaticasDANIEL ALEJANDRONessuna valutazione finora
- Anatomia Aplicada 11º Sistema Nervioso Generalidades Parte 1Documento25 pagineAnatomia Aplicada 11º Sistema Nervioso Generalidades Parte 1DANIEL ALEJANDRO100% (2)
- Muñeca y ManoDocumento22 pagineMuñeca y ManoDANIEL ALEJANDRONessuna valutazione finora
- Objetivos Formas Detrabajo en Spining para Gente de 40-50 AñosDocumento18 pagineObjetivos Formas Detrabajo en Spining para Gente de 40-50 AñosDANIEL ALEJANDRONessuna valutazione finora
- METABOLISMO PowerDocumento22 pagineMETABOLISMO PowerDANIEL ALEJANDRONessuna valutazione finora
- METABOLISMODocumento10 pagineMETABOLISMODANIEL ALEJANDRONessuna valutazione finora
- Valoración de La FuerzaDocumento14 pagineValoración de La FuerzaDANIEL ALEJANDRONessuna valutazione finora
- MuscularDocumento21 pagineMuscularDANIEL ALEJANDRO100% (1)
- Lesiones Oseas y Meniscales de La Rodillla 2da ParteDocumento15 pagineLesiones Oseas y Meniscales de La Rodillla 2da ParteDANIEL ALEJANDRONessuna valutazione finora
- Ejercicios Hipotiroidismo PDFDocumento14 pagineEjercicios Hipotiroidismo PDFDANIEL ALEJANDRO75% (4)
- Fisiologia Bioquimica JMCiordiaDocumento24 pagineFisiologia Bioquimica JMCiordiaDANIEL ALEJANDRONessuna valutazione finora
- Índice SanitatiaDocumento5 pagineÍndice SanitatiaDANIEL ALEJANDRONessuna valutazione finora
- Enfermedad de ParkisonDocumento20 pagineEnfermedad de ParkisonDANIEL ALEJANDRONessuna valutazione finora
- Enfermedades CardiovascularesDocumento19 pagineEnfermedades CardiovascularesDANIEL ALEJANDRONessuna valutazione finora
- EscoliosisDocumento19 pagineEscoliosisDANIEL ALEJANDRO100% (1)
- Educación Física en Niños Con TdahDocumento17 pagineEducación Física en Niños Con TdahDANIEL ALEJANDRONessuna valutazione finora
- Dolor PosturalDocumento10 pagineDolor PosturalDANIEL ALEJANDRONessuna valutazione finora
- Dolor PosturalDocumento17 pagineDolor PosturalDANIEL ALEJANDRONessuna valutazione finora
- Esquema Corporal BiomDocumento26 pagineEsquema Corporal BiomDANIEL ALEJANDRONessuna valutazione finora
- Ejercicios Hipotiroidismo PDFDocumento14 pagineEjercicios Hipotiroidismo PDFDANIEL ALEJANDRO75% (4)
- Sanitaria TPDocumento64 pagineSanitaria TPDANIEL ALEJANDRONessuna valutazione finora
- TP Uai 2Documento28 pagineTP Uai 2DANIEL ALEJANDRONessuna valutazione finora
- Presentacion 2Documento14 paginePresentacion 2DANIEL ALEJANDRONessuna valutazione finora
- Presentación 1Documento15 paginePresentación 1DANIEL ALEJANDRONessuna valutazione finora
- Presentacion 3Documento18 paginePresentacion 3DANIEL ALEJANDRONessuna valutazione finora
- Semana 3Documento33 pagineSemana 3Alexis Cliverth Briceño MorilloNessuna valutazione finora
- Aparato LocomotorDocumento44 pagineAparato Locomotorsanchezgalan100% (3)
- 01 Matriz CitoplasmaticaDocumento34 pagine01 Matriz CitoplasmaticaBill GeorgeNessuna valutazione finora
- FISIOLOGIA GUYTON UNIDAD 1 SEMANA 2 Capitulo 6Documento2 pagineFISIOLOGIA GUYTON UNIDAD 1 SEMANA 2 Capitulo 6LUCIA DEL CARMEN OTOYA CORONELNessuna valutazione finora
- Bromatologia de La CarneDocumento40 pagineBromatologia de La CarneHelios Villegas MezaNessuna valutazione finora
- Anatomia. Tarea 3 PDFDocumento22 pagineAnatomia. Tarea 3 PDFLuis DominguezNessuna valutazione finora
- Musculo Esqueletico Anatomia FisiologiaDocumento47 pagineMusculo Esqueletico Anatomia FisiologiacarolinaNessuna valutazione finora
- Banco de Preguntas Bio-8-IiDocumento3 pagineBanco de Preguntas Bio-8-IiDgh LggNessuna valutazione finora
- Bioquìmica de La Contracciòn MuscularDocumento4 pagineBioquìmica de La Contracciòn MuscularNicole Tupayachi ArceNessuna valutazione finora
- Tejido MuscularDocumento9 pagineTejido MuscularOscar PerezNessuna valutazione finora
- Contracción MuscularDocumento26 pagineContracción MuscularFabricio MartinezNessuna valutazione finora
- Fisiología de La Membrana, El Nervio y El Músculo.Documento14 pagineFisiología de La Membrana, El Nervio y El Músculo.Gabriel GuedezNessuna valutazione finora
- Clase 5Documento98 pagineClase 5Boo3010Nessuna valutazione finora
- ManualPLUS CARDioful 2017 CP PDFDocumento200 pagineManualPLUS CARDioful 2017 CP PDFAnonymous alzR2b1pq100% (1)
- Organelos Fibra MuscularDocumento5 pagineOrganelos Fibra Muscularyukitha88% (8)
- Medicina - Fisiología. Placa NeuromuscularDocumento5 pagineMedicina - Fisiología. Placa Neuromuscularmariaprincesita0% (1)
- MúsculoDocumento1 paginaMúsculoLaimaDelmarFonsecaCastellanosNessuna valutazione finora
- Nejmra Parturition - En.esDocumento13 pagineNejmra Parturition - En.esMarce Alvarado cmarcelaasNessuna valutazione finora
- 2 Potencial de Membrana y Contraccion MuscularDocumento16 pagine2 Potencial de Membrana y Contraccion MuscularFernando Caro MenaNessuna valutazione finora
- Trabajo de Grado UnicatolicaDocumento83 pagineTrabajo de Grado UnicatolicaPablo Cuevas SaldivarNessuna valutazione finora
- Tipeo de Pescado, Carne y Bioquimica PostcosechaDocumento109 pagineTipeo de Pescado, Carne y Bioquimica PostcosechaFelipe Antonio SalvadøNessuna valutazione finora
- Clasificación Del Tejido MuscularDocumento13 pagineClasificación Del Tejido MuscularJUAN CARLOS ROBAYO BOTIVA72% (18)
- Reflejos, Musculo Liso, Motoneurona Alfa y GammaDocumento3 pagineReflejos, Musculo Liso, Motoneurona Alfa y Gammajose araus jimenezNessuna valutazione finora
- FISIOLOGÍA 1 TRABAJO 3 - AndreaDocumento8 pagineFISIOLOGÍA 1 TRABAJO 3 - AndreaSUSAN ANDREA DORADONessuna valutazione finora
- Bioquímica de La CarneDocumento62 pagineBioquímica de La CarneCatalina HormazábalNessuna valutazione finora




![<![CDATA[Semiología neurológica]]>](https://imgv2-1-f.scribdassets.com/img/word_document/641540652/149x198/48e836a3e7/1714758029?v=1)