Potrebbero piacerti anche
- Seeing Eye To Eye: Vertebrate Eye Evolution and Adaptive RadiationDocumento7 pagineSeeing Eye To Eye: Vertebrate Eye Evolution and Adaptive RadiationAprille Octaviano-AsiloNessuna valutazione finora
- Vampire NumberDocumento3 pagineVampire NumberВук ДражићNessuna valutazione finora
- PoultryAnatomyEng WDocumento1 paginaPoultryAnatomyEng WBryan NicollNessuna valutazione finora
- Pigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingDa EverandPigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingNessuna valutazione finora
- Bird VisionDocumento13 pagineBird VisionХристинаГулеваNessuna valutazione finora
- Exhibit PDFDocumento15 pagineExhibit PDFJaffer RazaNessuna valutazione finora
- A Birds Eye View of The Aimst Uni (7 March 2015)Documento174 pagineA Birds Eye View of The Aimst Uni (7 March 2015)M.Ravichandran100% (1)
- Mammals PDFDocumento2 pagineMammals PDFron noeNessuna valutazione finora
- Available Plant Unit ResourcesDocumento5 pagineAvailable Plant Unit Resourcesapi-280716018Nessuna valutazione finora
- By Diego and Hector. 5th ADocumento6 pagineBy Diego and Hector. 5th AangelcealoNessuna valutazione finora
- Anatomy (Stockoe) For Mini NotesDocumento86 pagineAnatomy (Stockoe) For Mini NotesIvy Dianne PascualNessuna valutazione finora
- Head and Tails Animals PDFDocumento7 pagineHead and Tails Animals PDFOtilia PavalNessuna valutazione finora
- Swallowtail ButterflyDocumento8 pagineSwallowtail Butterflymineiro_quimica2007Nessuna valutazione finora
- Disection of A PigeonDocumento62 pagineDisection of A PigeonErnani BatimanaNessuna valutazione finora
- Laughing KookaburraDocumento3 pagineLaughing KookaburraDbaltNessuna valutazione finora
- A History of EntomologicalDocumento20 pagineA History of EntomologicalBarzola HerlissNessuna valutazione finora
- Snakes On An Enrichment Programme?!: Shelby Stanfield, MSC James Brereton, MSCDocumento11 pagineSnakes On An Enrichment Programme?!: Shelby Stanfield, MSC James Brereton, MSCWarner LlaveNessuna valutazione finora
- Class AvesDocumento11 pagineClass AveshemavarshiniNessuna valutazione finora
- Reptile ReproductionDocumento5 pagineReptile ReproductionDiyar Ahmad100% (1)
- Raptor Nutrition enDocumento39 pagineRaptor Nutrition enMark AbellaNessuna valutazione finora
- Internal Anatomy: Skeletal SystemDocumento6 pagineInternal Anatomy: Skeletal Systemroyleewheels100% (1)
- RAP Example Lessons CombinedDocumento52 pagineRAP Example Lessons CombinedSeema ChauhanNessuna valutazione finora
- 2003 Avian Care Manual PDFDocumento132 pagine2003 Avian Care Manual PDFRonald ChavezNessuna valutazione finora
- 08 Animal Behavior Restraint Equine HANDOUTDocumento2 pagine08 Animal Behavior Restraint Equine HANDOUTJonathan W. VillacísNessuna valutazione finora
- Bird Anatomy: From Wikipedia, The Free EncyclopediaDocumento12 pagineBird Anatomy: From Wikipedia, The Free Encyclopediahd_catwolfieNessuna valutazione finora
- TriviumDocumento231 pagineTriviumAther Abdul JabbarNessuna valutazione finora
- Penguin Husbandry Manual Third Edition 2005Documento142 paginePenguin Husbandry Manual Third Edition 2005westleybatistaNessuna valutazione finora
- General Characters of Class AvesDocumento10 pagineGeneral Characters of Class AvesGEETA MOHAN100% (3)
- Feather Destructive BehaviorDocumento20 pagineFeather Destructive BehaviorAnh-MinhNessuna valutazione finora
- PeacockDocumento12 paginePeacockparag03Nessuna valutazione finora
- Pigeon Breeds All List NPADocumento2 paginePigeon Breeds All List NPApigeontypeNessuna valutazione finora
- Animal Behaviour & Ethology Avneesh BishnoiDocumento20 pagineAnimal Behaviour & Ethology Avneesh BishnoiAishwary SinghNessuna valutazione finora
- Common OstrichDocumento165 pagineCommon OstrichHarish BhatNessuna valutazione finora
- Cockatoo EnrichmentDocumento10 pagineCockatoo EnrichmentOmary2jNessuna valutazione finora
- 1 2019 Herpetology PPTDocumento45 pagine1 2019 Herpetology PPTBryceNessuna valutazione finora
- Horse BehaviorDocumento2 pagineHorse BehaviormmadomadNessuna valutazione finora
- Inflammation AvianDocumento4 pagineInflammation Avianamit vishen75% (4)
- Hornbill Husbandry Manual (EAZA)Documento67 pagineHornbill Husbandry Manual (EAZA)Lâm Kim HảiNessuna valutazione finora
- Surgical Anatomy Avian, Thoracic and Hind Limbs (VetBooks - Ir)Documento83 pagineSurgical Anatomy Avian, Thoracic and Hind Limbs (VetBooks - Ir)LorrayneNessuna valutazione finora
- Galah CockatooDocumento2 pagineGalah CockatooColin LewisNessuna valutazione finora
- Animal TrainingDocumento44 pagineAnimal TrainingMunir Yusufzai100% (1)
- Reptile EmergencyDocumento16 pagineReptile EmergencyPedro Trotsky DeathMaskNessuna valutazione finora
- 39 Canaries Finches MynahsDocumento36 pagine39 Canaries Finches Mynahsadel malikNessuna valutazione finora
- Mammals ppt-1Documento10 pagineMammals ppt-1api-309894157Nessuna valutazione finora
- Sow Body Condition Scoring Guidelines: Condición Corporal 1Documento1 paginaSow Body Condition Scoring Guidelines: Condición Corporal 1imonetaNessuna valutazione finora
- Asset - 8288 - Anatomy of The Canine Forelimb PDFDocumento35 pagineAsset - 8288 - Anatomy of The Canine Forelimb PDFBianca ElenaNessuna valutazione finora
- Mammals That Have Wings and Can FlyDocumento3 pagineMammals That Have Wings and Can FlyrapaseNessuna valutazione finora
- Kestrels of The World 11-11-11Documento48 pagineKestrels of The World 11-11-11DelphinitaNessuna valutazione finora
- ANATOMY OF PIGEON (Columba Domestica)Documento12 pagineANATOMY OF PIGEON (Columba Domestica)Mellya RizkiNessuna valutazione finora
- Handbook of The Birds of India and Pakistan V 3Documento367 pagineHandbook of The Birds of India and Pakistan V 3Ghaier KazmiNessuna valutazione finora
- The Mirror of Literature, Amusement, and Instruction Volume 19, No. 550, June 2, 1832 by VariousDocumento30 pagineThe Mirror of Literature, Amusement, and Instruction Volume 19, No. 550, June 2, 1832 by VariousGutenberg.org100% (1)
- General Characters of Class ReptiliaDocumento20 pagineGeneral Characters of Class ReptiliaErin Martinez100% (4)
- UltrasonographyDocumento21 pagineUltrasonographyChecko LatteNessuna valutazione finora
- Haginbalds Arte of Medieval FaulconryDocumento23 pagineHaginbalds Arte of Medieval FaulconryimmortalskyNessuna valutazione finora
- Characteristics of Class AvesDocumento4 pagineCharacteristics of Class AvesPriti Ranjan NarendraNessuna valutazione finora
- Domestication of The HorseDocumento11 pagineDomestication of The HorseMuhammad Farrukh Hafeez100% (1)
- Care Sheet - Amethystine or Scrub Python (Morelia Amethistina)Documento2 pagineCare Sheet - Amethystine or Scrub Python (Morelia Amethistina)John Gamesby100% (1)
- Ayam Kampung PDFDocumento9 pagineAyam Kampung PDFmedeNessuna valutazione finora
- 181 181 1 PBDocumento7 pagine181 181 1 PBHi KittyNessuna valutazione finora
- Burung Puyuh PDFDocumento7 pagineBurung Puyuh PDFmedeNessuna valutazione finora
- Kelinci PDFDocumento11 pagineKelinci PDFmedeNessuna valutazione finora
- Itik PDFDocumento7 pagineItik PDFmedeNessuna valutazione finora
- Babi PDFDocumento15 pagineBabi PDFmedeNessuna valutazione finora
- Babi PDFDocumento15 pagineBabi PDFmedeNessuna valutazione finora
- Itik PDFDocumento7 pagineItik PDFmedeNessuna valutazione finora
- Layer PDFDocumento9 pagineLayer PDFmedeNessuna valutazione finora
- Layer PDFDocumento9 pagineLayer PDFmedeNessuna valutazione finora
- Ayam Kampung PDFDocumento9 pagineAyam Kampung PDFmedeNessuna valutazione finora
- Kualitas Fisik Dan Nutrisi Jerami Padi Fermentasi Pada Berbagai Penambahan StarterDocumento10 pagineKualitas Fisik Dan Nutrisi Jerami Padi Fermentasi Pada Berbagai Penambahan StartermedeNessuna valutazione finora
- Peerj 4634 PDFDocumento11 paginePeerj 4634 PDFShilpa KßhNessuna valutazione finora
- Peerj 4634 PDFDocumento11 paginePeerj 4634 PDFShilpa KßhNessuna valutazione finora
- Burung Puyuh PDFDocumento7 pagineBurung Puyuh PDFmedeNessuna valutazione finora
- Burung Puyuh PDFDocumento7 pagineBurung Puyuh PDFmedeNessuna valutazione finora
- Kelinci PDFDocumento11 pagineKelinci PDFmedeNessuna valutazione finora
- Kelinci PDFDocumento11 pagineKelinci PDFmedeNessuna valutazione finora
- Babi PDFDocumento15 pagineBabi PDFmedeNessuna valutazione finora
- Peerj 4634 PDFDocumento11 paginePeerj 4634 PDFShilpa KßhNessuna valutazione finora
- Ayam Kampung PDFDocumento9 pagineAyam Kampung PDFmedeNessuna valutazione finora
- Peerj 4634 PDFDocumento11 paginePeerj 4634 PDFShilpa KßhNessuna valutazione finora
- Itik PDFDocumento7 pagineItik PDFmedeNessuna valutazione finora
- Layer PDFDocumento9 pagineLayer PDFmedeNessuna valutazione finora
- Genomic Regions Influencing Intramuscular Fat in Divergently Selected Rabbit LinesDocumento12 pagineGenomic Regions Influencing Intramuscular Fat in Divergently Selected Rabbit LinesmedeNessuna valutazione finora
- Geographic Morphological Variation of Gentoo Penguin (Pygoscelis Papua) and Sex Identification: Using Morphometric Characters and Molecular MarkersDocumento13 pagineGeographic Morphological Variation of Gentoo Penguin (Pygoscelis Papua) and Sex Identification: Using Morphometric Characters and Molecular MarkersmedeNessuna valutazione finora
- PenguinsDocumento4 paginePenguinsTaylur CalandraNessuna valutazione finora
- PenguinsDocumento4 paginePenguinsTaylur CalandraNessuna valutazione finora
- Biology of PenguinsDocumento43 pagineBiology of PenguinsMarilyn Castillo100% (10)
- Corpsman Pt1Documento140 pagineCorpsman Pt1David Radcliffe100% (1)
- Nur 194 - Care of Mother, Child, & AdolescentDocumento43 pagineNur 194 - Care of Mother, Child, & AdolescentAileen Cagulada100% (1)
- OEB 139 Adams Lin Zhang Final Paper DraftDocumento7 pagineOEB 139 Adams Lin Zhang Final Paper Draftd-fbuser-35748307Nessuna valutazione finora
- Lossing BerylArbucklesCranialDocumento67 pagineLossing BerylArbucklesCranialUlisses Rodrigues Da Silva100% (1)
- The Nursing Care of A Family With A NewbornDocumento5 pagineThe Nursing Care of A Family With A NewbornDonell EscalanteNessuna valutazione finora
- Sistem Rangka (Anatomi) - DikonversiDocumento17 pagineSistem Rangka (Anatomi) - Dikonversinitstay325Nessuna valutazione finora
- ENT RadiologiDocumento70 pagineENT RadiologiyowwwNessuna valutazione finora
- Axial Skeleton, ENDOSKELETON OF VERTEBRATESDocumento52 pagineAxial Skeleton, ENDOSKELETON OF VERTEBRATESDave MarimonNessuna valutazione finora
- The Skeleton: Chapter 7 - Part ADocumento7 pagineThe Skeleton: Chapter 7 - Part AJonathan HigginbothamNessuna valutazione finora
- Nissin Tooth Anatomy AreaDocumento6 pagineNissin Tooth Anatomy AreaDanis Diba Sabatillah YaminNessuna valutazione finora
- Rapid Maxillary ExpansionDocumento55 pagineRapid Maxillary ExpansionDon George GeojanNessuna valutazione finora
- Bush CV 7 8 15Documento20 pagineBush CV 7 8 15api-296497193Nessuna valutazione finora
- Skull Procedures 1. PA Projection 2. AP ProjectionDocumento215 pagineSkull Procedures 1. PA Projection 2. AP ProjectionradioNessuna valutazione finora
- sKuLL Positioning sTuDy GuiDe!!!Documento6 paginesKuLL Positioning sTuDy GuiDe!!!Ling YuNessuna valutazione finora
- The Mewing BibleDocumento13 pagineThe Mewing BibledudusancavaknewNessuna valutazione finora
- Sexing The Human SkullDocumento5 pagineSexing The Human SkullAisyah RieskiuNessuna valutazione finora
- Compensatory MechanismDocumento30 pagineCompensatory MechanismSenthil Kumar Elumalai100% (1)
- Laboratory 7Documento8 pagineLaboratory 7Diana Rose SaludoNessuna valutazione finora
- Snakes Found in IndiaDocumento246 pagineSnakes Found in IndiaBalu SamyNessuna valutazione finora
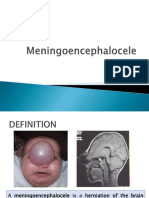
- MeningoencephaloceleDocumento13 pagineMeningoencephaloceleIda Bagus Yudha Prasista100% (1)
- Craniofacial Osteology 1Documento36 pagineCraniofacial Osteology 1Yeow Yun ChihNessuna valutazione finora
- Vinca Belo BrdoDocumento25 pagineVinca Belo BrdoДимитрије МарковићNessuna valutazione finora
- 5-Locomotion and Movement PDFDocumento4 pagine5-Locomotion and Movement PDFPraveen KumarNessuna valutazione finora
- Growth and Development of Mandible / Orthodontic Courses by Indian Dental AcademyDocumento78 pagineGrowth and Development of Mandible / Orthodontic Courses by Indian Dental Academyindian dental academyNessuna valutazione finora
- Deformational Plagiocephaly Brachycephaly And.4Documento8 pagineDeformational Plagiocephaly Brachycephaly And.4Neith Velez PachecoNessuna valutazione finora
- TNCC StudyDocumento22 pagineTNCC StudyDallas Lindsey100% (5)
- Cambridge Primary Science Learner's Book Stage 4 - Public PDFDocumento13 pagineCambridge Primary Science Learner's Book Stage 4 - Public PDFDavid Musila Toywa100% (3)
- Semana 2 (2) Park (2005)Documento5 pagineSemana 2 (2) Park (2005)Arturo SanabriaNessuna valutazione finora
- AAB Human Skull Anatomy ActivityDocumento2 pagineAAB Human Skull Anatomy ActivityCamila VorkapicNessuna valutazione finora
- Science Demo q2 LP (Ibl, 5e's, Cai With Di) by MebDocumento6 pagineScience Demo q2 LP (Ibl, 5e's, Cai With Di) by MebDORY WINGNessuna valutazione finora
- Why We Die: The New Science of Aging and the Quest for ImmortalityDa EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityValutazione: 4 su 5 stelle4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDa EverandTales from Both Sides of the Brain: A Life in NeuroscienceValutazione: 3 su 5 stelle3/5 (18)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDa EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisValutazione: 3.5 su 5 stelle3.5/5 (2)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDa EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsValutazione: 4 su 5 stelle4/5 (5)
- Gut: the new and revised Sunday Times bestsellerDa EverandGut: the new and revised Sunday Times bestsellerValutazione: 4 su 5 stelle4/5 (392)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDa EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedValutazione: 4 su 5 stelle4/5 (11)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDa EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeNessuna valutazione finora
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Da EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Valutazione: 4 su 5 stelle4/5 (378)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDa Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessValutazione: 4 su 5 stelle4/5 (33)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDa EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperValutazione: 4.5 su 5 stelle4.5/5 (15)
- Seven and a Half Lessons About the BrainDa EverandSeven and a Half Lessons About the BrainValutazione: 4 su 5 stelle4/5 (109)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDa EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouValutazione: 4.5 su 5 stelle4.5/5 (62)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDa EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceValutazione: 4.5 su 5 stelle4.5/5 (516)
- Human: The Science Behind What Makes Your Brain UniqueDa EverandHuman: The Science Behind What Makes Your Brain UniqueValutazione: 3.5 su 5 stelle3.5/5 (38)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDa EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionValutazione: 4 su 5 stelle4/5 (811)
- Good Without God: What a Billion Nonreligious People Do BelieveDa EverandGood Without God: What a Billion Nonreligious People Do BelieveValutazione: 4 su 5 stelle4/5 (66)
- The Invention of Tomorrow: A Natural History of ForesightDa EverandThe Invention of Tomorrow: A Natural History of ForesightValutazione: 4.5 su 5 stelle4.5/5 (5)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildDa EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildValutazione: 4.5 su 5 stelle4.5/5 (44)
- Who's in Charge?: Free Will and the Science of the BrainDa EverandWho's in Charge?: Free Will and the Science of the BrainValutazione: 4 su 5 stelle4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDa EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNessuna valutazione finora
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDa EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesValutazione: 4.5 su 5 stelle4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDa EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldValutazione: 4.5 su 5 stelle4.5/5 (18)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessDa EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessValutazione: 4 su 5 stelle4/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDa EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondValutazione: 4 su 5 stelle4/5 (4)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDa EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemValutazione: 4.5 su 5 stelle4.5/5 (115)
- Lymph & Longevity: The Untapped Secret to HealthDa EverandLymph & Longevity: The Untapped Secret to HealthValutazione: 4.5 su 5 stelle4.5/5 (13)