Potrebbero piacerti anche
- Niveles de Organización de Los Seres VivosDocumento11 pagineNiveles de Organización de Los Seres VivosLudamour Javela Másmela100% (1)
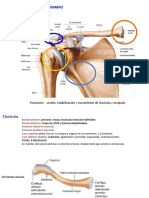
- Clase 2 - Hombro 2019Documento20 pagineClase 2 - Hombro 2019Yébeny Tufino AlvaradoNessuna valutazione finora
- Ontogenia Linfocitos BDocumento57 pagineOntogenia Linfocitos BXimena Granados MuñozNessuna valutazione finora
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Da EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Nessuna valutazione finora
- Ontogenia Linfocitos BDocumento3 pagineOntogenia Linfocitos BIna GataNessuna valutazione finora
- Inmunidad HumoralDocumento36 pagineInmunidad HumoralLesly Elizabeth Tacanga EspinozaNessuna valutazione finora
- Hidrolisis de RNA y DNA e Identificación de Bases Púricas y Pirimídicas Por Cromatografía en Capa Fina.Documento2 pagineHidrolisis de RNA y DNA e Identificación de Bases Púricas y Pirimídicas Por Cromatografía en Capa Fina.Monse Blanquel25% (4)
- Biol emDocumento99 pagineBiol emJoel VegaNessuna valutazione finora
- URP Guía de Prácticas Microbiologia 2020 - 1Documento82 pagineURP Guía de Prácticas Microbiologia 2020 - 1Yébeny Tufino Alvarado100% (1)
- Inmunogenética 2,015Documento60 pagineInmunogenética 2,015Brisa NarvaezNessuna valutazione finora
- Anticuerpos PDFDocumento4 pagineAnticuerpos PDFEmil OCNessuna valutazione finora
- ANTICUERPOSDocumento4 pagineANTICUERPOSOctavio ZmNessuna valutazione finora
- InmunoglobulinasDocumento30 pagineInmunoglobulinasTifoneitorNessuna valutazione finora
- Activacion de Los Linfocito B Resumen Parcial 2 InmunoDocumento14 pagineActivacion de Los Linfocito B Resumen Parcial 2 InmunoFrank AlexanderNessuna valutazione finora
- Estructura y Funcion de Los AnticuerposDocumento3 pagineEstructura y Funcion de Los Anticuerposlucyzoe_2805Nessuna valutazione finora
- InmunoglobulinaDocumento16 pagineInmunoglobulinaVerónica CisnerosNessuna valutazione finora
- M InmunologiaDocumento126 pagineM InmunologiaHilma Bermudez Gonzalez100% (1)
- Cap 4. Reconocimiento Del Antígeno en El Sistema Inmunitario AdaptativoDocumento11 pagineCap 4. Reconocimiento Del Antígeno en El Sistema Inmunitario AdaptativoSebastián FranciscoNessuna valutazione finora
- 2T Receptores BCR y TCRDocumento22 pagine2T Receptores BCR y TCRPaula Gil AlvarezNessuna valutazione finora
- Linfocitos BDocumento38 pagineLinfocitos Blady laura restrepoNessuna valutazione finora
- Clase de Antígenos, AnticuerposDocumento48 pagineClase de Antígenos, AnticuerposFranciscoA.BustamanteNessuna valutazione finora
- Receptores de Los LsDocumento11 pagineReceptores de Los LsPaola PintoNessuna valutazione finora
- Ontogenia B. El Delicado Equilibrio Entre La Diversidad y La AutoinmundadDocumento12 pagineOntogenia B. El Delicado Equilibrio Entre La Diversidad y La AutoinmundadDiana GarcíaNessuna valutazione finora
- Anticuerpo y AntigenoDocumento24 pagineAnticuerpo y Antigenomajo gsNessuna valutazione finora
- Diversidad de Las InmunoglobulinasDocumento7 pagineDiversidad de Las InmunoglobulinasladbymNessuna valutazione finora
- Desarrollo de Los Linfocitos B ResumenDocumento12 pagineDesarrollo de Los Linfocitos B ResumenJGRBBKDVRNessuna valutazione finora
- Clase 8 - Ontogenia BDocumento10 pagineClase 8 - Ontogenia BLautaro VasqueNessuna valutazione finora
- Taller Bioquimica 3 AlexaDocumento2 pagineTaller Bioquimica 3 AlexaAlexandra Niño BeltranNessuna valutazione finora
- 10 Inmunoglobulinas e Inmunidad HumoralDocumento27 pagine10 Inmunoglobulinas e Inmunidad HumoralJ. JoseNessuna valutazione finora
- 4.2.1 Cascadas de Señalización Por El BCR y Correceptor Del Linfocito BDocumento8 pagine4.2.1 Cascadas de Señalización Por El BCR y Correceptor Del Linfocito Bgarmyoftomorrow103Nessuna valutazione finora
- Activación Linfocitos B1Documento33 pagineActivación Linfocitos B1Alejandro Josue Leon MedinaNessuna valutazione finora
- AnticuerposDocumento58 pagineAnticuerposnomade.astralNessuna valutazione finora
- Capitulo 10 InmunologiaDocumento9 pagineCapitulo 10 InmunologiaDayanara PlazaNessuna valutazione finora
- Resumen Documento Linfocitos BDocumento10 pagineResumen Documento Linfocitos BYesenia Aracely Juarez PopNessuna valutazione finora
- Linfocitos Ensayo de Inmuno PDFDocumento6 pagineLinfocitos Ensayo de Inmuno PDFNicole Burgos VeraNessuna valutazione finora
- Genética de Las IgsDocumento7 pagineGenética de Las IgsFreddy TorresNessuna valutazione finora
- Bloque 4 Preguntas - Adrian Muñoz CastroDocumento6 pagineBloque 4 Preguntas - Adrian Muñoz CastroAdrian MuñozNessuna valutazione finora
- PatologíaDocumento14 paginePatologíaVivian CárdenasNessuna valutazione finora
- Desarrollo y Maduración de Los LinfocitosDocumento66 pagineDesarrollo y Maduración de Los LinfocitosLeonardo LeonNessuna valutazione finora
- CAPITULO Inmunidad Mediada Por Linfocitos BDocumento15 pagineCAPITULO Inmunidad Mediada Por Linfocitos BLUCAS NICOLAS MASINNessuna valutazione finora
- Inmunoglobulinas y BCRDocumento14 pagineInmunoglobulinas y BCRLucas Francisco Vargas RodriguezNessuna valutazione finora
- Cuestionario N°3 AnticuerposDocumento5 pagineCuestionario N°3 AnticuerposMaria RadcliffeNessuna valutazione finora
- InmunogDocumento11 pagineInmunogcarosolanoNessuna valutazione finora
- Anticuerpos o InmunoglobulinasDocumento8 pagineAnticuerpos o InmunoglobulinasL PDNessuna valutazione finora
- Tiraje 8 - Anticuerpos. Estructura y FuncionDocumento12 pagineTiraje 8 - Anticuerpos. Estructura y FuncionGabriela GuevaraNessuna valutazione finora
- Repaso Capitulo 5-9Documento10 pagineRepaso Capitulo 5-9Araceli hernandez JaimeNessuna valutazione finora
- 09 LinfocitosDocumento38 pagine09 LinfocitosdraanaisysNessuna valutazione finora
- 7 Clase Inmuinologia Upsjb 2017 1Documento15 pagine7 Clase Inmuinologia Upsjb 2017 1JholcarNessuna valutazione finora
- Antigen OsDocumento4 pagineAntigen OsDiana ArmasNessuna valutazione finora
- TEMA 5. Anticuerpos y BCRDocumento20 pagineTEMA 5. Anticuerpos y BCRNina Souto BlancoNessuna valutazione finora
- INMUNOGLOBULINASDocumento5 pagineINMUNOGLOBULINASLizbthNessuna valutazione finora
- Inmunoglobulina eDocumento26 pagineInmunoglobulina eLpMr Marco AntonioNessuna valutazione finora
- Antigenos y AnticuerposDocumento34 pagineAntigenos y AnticuerposmariaNessuna valutazione finora
- Receptores Específicos de Antígeno de Los Linfocitos T y BDocumento8 pagineReceptores Específicos de Antígeno de Los Linfocitos T y BPaulina MayaNessuna valutazione finora
- TEMA 1 InmunoDocumento13 pagineTEMA 1 InmunoGabriel Hernandez MartinNessuna valutazione finora
- Clase 004b INMUNOLOGIA Activación Linfocitos B InmunoglobulinasDocumento71 pagineClase 004b INMUNOLOGIA Activación Linfocitos B InmunoglobulinasFernando Davila valleNessuna valutazione finora
- Clase 7 - La Respuesta Inmunitaria Humoral y El Sistema de ComplementoDocumento61 pagineClase 7 - La Respuesta Inmunitaria Humoral y El Sistema de Complementocory ramirezNessuna valutazione finora
- Inmunidad Mediada Por Linfocitos BDocumento85 pagineInmunidad Mediada Por Linfocitos Bjustkim1903Nessuna valutazione finora
- Inmuno-Proliferacion Clonal de Los LinfocitosDocumento4 pagineInmuno-Proliferacion Clonal de Los LinfocitosIsrael CabreraNessuna valutazione finora
- Anticuerpo CREDocumento8 pagineAnticuerpo CRERec 379Nessuna valutazione finora
- LLA-B Parte 2: Ontogenia de Linfocitos T y BDocumento14 pagineLLA-B Parte 2: Ontogenia de Linfocitos T y BEri MñzNessuna valutazione finora
- INMUNOGENETICADocumento63 pagineINMUNOGENETICAFrank García Rojas0% (1)
- Aspectos Moleculares y Funcionales de Los AnticuerposDocumento2 pagineAspectos Moleculares y Funcionales de Los AnticuerposDiego Castaño MejiaNessuna valutazione finora
- LB Otras Poblaciones de LBDocumento4 pagineLB Otras Poblaciones de LBBioqui En ProcesoNessuna valutazione finora
- Resumen Clase 5Documento4 pagineResumen Clase 5Dayanna MorenoNessuna valutazione finora
- URP Seminario 01 Filogenia Del Sistema InmuneDocumento25 pagineURP Seminario 01 Filogenia Del Sistema InmuneYébeny Tufino AlvaradoNessuna valutazione finora
- Clase 5. Artrópodos I-ArácnidosDocumento33 pagineClase 5. Artrópodos I-ArácnidosYébeny Tufino AlvaradoNessuna valutazione finora
- URP Seminario 03 Receptores SolublesDocumento13 pagineURP Seminario 03 Receptores SolublesYébeny Tufino AlvaradoNessuna valutazione finora
- Informe 11Documento2 pagineInforme 11Yébeny Tufino AlvaradoNessuna valutazione finora
- Clase 2. Virología Básica. JHVP. 2017.1Documento14 pagineClase 2. Virología Básica. JHVP. 2017.1Yébeny Tufino AlvaradoNessuna valutazione finora
- Guia Fisio 2020-IDocumento68 pagineGuia Fisio 2020-IYébeny Tufino AlvaradoNessuna valutazione finora
- Laboratorio 3Documento2 pagineLaboratorio 3Yébeny Tufino AlvaradoNessuna valutazione finora
- HEMATOODocumento10 pagineHEMATOOYébeny Tufino Alvarado100% (1)
- Semana 1 Estructura de La PielDocumento48 pagineSemana 1 Estructura de La PielYébeny Tufino AlvaradoNessuna valutazione finora
- Semana 2 Estructura de Los Anexos CutaneosDocumento50 pagineSemana 2 Estructura de Los Anexos CutaneosYébeny Tufino AlvaradoNessuna valutazione finora
- Cuestionario - Lab 4Documento4 pagineCuestionario - Lab 4Yébeny Tufino AlvaradoNessuna valutazione finora
- Cuestionario - Lab 5Documento2 pagineCuestionario - Lab 5Yébeny Tufino Alvarado0% (3)
- FosfolípidosDocumento3 pagineFosfolípidosFräncïscö MëjíäNessuna valutazione finora
- Silabo BiologiaDocumento9 pagineSilabo BiologiaCAROL YAMILY RAMIREZ CUEVANessuna valutazione finora
- Bio t17 MutacionesDocumento5 pagineBio t17 MutacionesJason DarkussNessuna valutazione finora
- Qué Es La PurinaDocumento2 pagineQué Es La PurinaMaycon Alves100% (1)
- Celula Concepto y ClasificaciónDocumento2 pagineCelula Concepto y ClasificaciónSebastian Sanchez sssNessuna valutazione finora
- Clasificación de C, LyPDocumento23 pagineClasificación de C, LyPDorian Adriano CadenaNessuna valutazione finora
- Organelos Endomembranosos y No MembranososDocumento52 pagineOrganelos Endomembranosos y No MembranososmilenkaNessuna valutazione finora
- Tema #5 El Maravilloso Mundo de La CélulaDocumento4 pagineTema #5 El Maravilloso Mundo de La CélulaAbilio MontenegroNessuna valutazione finora
- Evaluacion Bmct-U07. Clonacion de Acidos Nucleicos.Documento8 pagineEvaluacion Bmct-U07. Clonacion de Acidos Nucleicos.ILimbus Nauticus100% (1)
- Glucogénesis y GlucogenólisisDocumento3 pagineGlucogénesis y GlucogenólisisDavidNessuna valutazione finora
- RetroviridaeDocumento41 pagineRetroviridaeMaria Alejandra Vargas QuinteroNessuna valutazione finora
- Universidad de Guadalajara: Centro Universitario de Ciencias Exactas e IngenieríasDocumento4 pagineUniversidad de Guadalajara: Centro Universitario de Ciencias Exactas e IngenieríasJORGE MORENO CERDANessuna valutazione finora
- Biologia - Clase1 - La Celula04Documento13 pagineBiologia - Clase1 - La Celula04sla_bmNessuna valutazione finora
- Examen 2 Morfo IDocumento2 pagineExamen 2 Morfo IRichard MatheusNessuna valutazione finora
- Paso 1 - Resolver El Cuestionario Sobre La Célula Sus Partes y Funciones - Biología AmbientalDocumento8 paginePaso 1 - Resolver El Cuestionario Sobre La Célula Sus Partes y Funciones - Biología AmbientalKatherine TorresNessuna valutazione finora
- Paper TraducidoDocumento6 paginePaper TraducidoMacarena Moreno FernándezNessuna valutazione finora
- Guia de BiologiaDocumento70 pagineGuia de BiologiaLucrecia LavarredaNessuna valutazione finora
- Aminoacidos y ProteinasDocumento20 pagineAminoacidos y ProteinasFran SolisNessuna valutazione finora
- Cucigrama Examen Dayana y WeimarDocumento2 pagineCucigrama Examen Dayana y WeimarDayana Garcés WatsteinNessuna valutazione finora
- 02 Transporte CelularDocumento5 pagine02 Transporte CelularBrenda KrausNessuna valutazione finora
- Examen Biología Extraordinaria de 2012Documento4 pagineExamen Biología Extraordinaria de 2012Mari Carmen Lopez GarciaNessuna valutazione finora
- Laboratorio 2023111816925 - 20231126 213453Documento1 paginaLaboratorio 2023111816925 - 20231126 213453mirandamatajudith331Nessuna valutazione finora
- Qué Son Los PrionesDocumento5 pagineQué Son Los Prionesyimilequimik100% (1)
- Examen Replicación Transcripción y Traducción 2º BNDocumento3 pagineExamen Replicación Transcripción y Traducción 2º BNVet SLNessuna valutazione finora
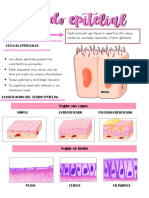
- Tejido EpitelialDocumento10 pagineTejido EpitelialAlessandra Sheyla Churata LlanosNessuna valutazione finora
- P9 TriacilgliceridosDocumento7 pagineP9 TriacilgliceridosElizabeth soto MontesNessuna valutazione finora
- SaviaDocumento3 pagineSaviahector vcNessuna valutazione finora