Potrebbero piacerti anche
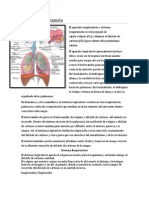
- Aparato respiratorio: anatomía y fisiologíaDocumento32 pagineAparato respiratorio: anatomía y fisiologíaSergio GalvanNessuna valutazione finora
- Alteraciones Del Sistema LinfáticoDocumento38 pagineAlteraciones Del Sistema LinfáticoDiana Roxana DorregarayNessuna valutazione finora
- Anexo Sistema RespiratorioDocumento5 pagineAnexo Sistema RespiratorioAlejandra OrellanaNessuna valutazione finora
- Af1-Mod 1-RespDocumento12 pagineAf1-Mod 1-RespGustavo CamaraNessuna valutazione finora
- Unidad Temática 4. Aparato RespiratorioDocumento11 pagineUnidad Temática 4. Aparato RespiratorioLara MayoralNessuna valutazione finora
- UNIDAD II: Cuidado de Las Personas Con Alteraciones en La Necesidad de Oxigenación.Documento6 pagineUNIDAD II: Cuidado de Las Personas Con Alteraciones en La Necesidad de Oxigenación.Daiana RojasNessuna valutazione finora
- Mecanica RespiratoriaDocumento9 pagineMecanica RespiratoriaRouslymar BolivarNessuna valutazione finora
- INVESTIGACIONDocumento14 pagineINVESTIGACIONTato GomesNessuna valutazione finora
- 14º Biofísica de La RespiraciónDocumento18 pagine14º Biofísica de La RespiraciónCarlos Hugo Graverolle LópezNessuna valutazione finora
- Documento Sin TítuloDocumento4 pagineDocumento Sin TítuloCarmen PeloNessuna valutazione finora
- Anatoma y Fisiologia Del Sistema RespiratorioDocumento4 pagineAnatoma y Fisiologia Del Sistema RespiratorioerickguzmanhuaytaNessuna valutazione finora
- Aparato RespiratorioDocumento2 pagineAparato RespiratorioJosemaNessuna valutazione finora
- Anatoma y Fisiologia Del Sistema RespiratorioDocumento5 pagineAnatoma y Fisiologia Del Sistema RespiratorioerickguzmanhuaytaNessuna valutazione finora
- Proyecto de AnatomiaDocumento13 pagineProyecto de AnatomiaKim LaoNessuna valutazione finora
- El intercambio de gases es la provisión de oxigeno de los pulmones al torrente sanguíneo y la eliminación del dióxido de carbono (CO2) del torrente sanguíneo a los pulmonesDocumento10 pagineEl intercambio de gases es la provisión de oxigeno de los pulmones al torrente sanguíneo y la eliminación del dióxido de carbono (CO2) del torrente sanguíneo a los pulmonesJose Ignacio HonoNessuna valutazione finora
- Sistema RespiratorioDocumento7 pagineSistema RespiratorioCalebAlccaNessuna valutazione finora
- Sistema RespiratorioDocumento10 pagineSistema RespiratorioNery TorresNessuna valutazione finora
- Sistema respiratorio y sus funcionesDocumento2 pagineSistema respiratorio y sus funcionesKevin CastilloNessuna valutazione finora
- Aparato RespiratorioDocumento6 pagineAparato RespiratorioCindy de los AngelesNessuna valutazione finora
- Aparato RespiratorioDocumento19 pagineAparato RespiratorioARMARLISNessuna valutazione finora
- Sistema Respiratorio HumanoDocumento7 pagineSistema Respiratorio HumanoMaria Jose Yazuri Condori AlanocaNessuna valutazione finora
- 18 - Sistema Respiratorio PDFDocumento4 pagine18 - Sistema Respiratorio PDFKike PaxekoNessuna valutazione finora
- Asignacion 1. RadiologiaDocumento20 pagineAsignacion 1. RadiologiaYorge Frías AcostaNessuna valutazione finora
- Sistema RespiratorioDocumento37 pagineSistema Respiratorioleydy lisbethNessuna valutazione finora
- El Sistema RespiratorioDocumento21 pagineEl Sistema RespiratorioOscar Andres Gonzalez GarciaNessuna valutazione finora
- InformeDocumento14 pagineInformeGustavo Tabo MamaniNessuna valutazione finora
- Aparato RespiratorioDocumento18 pagineAparato RespiratorioMalhi Mariana Rios Huarcaya0% (1)
- Anatomia Del Sistema RespiratorioDocumento6 pagineAnatomia Del Sistema RespiratorioAlix LopezNessuna valutazione finora
- Aparato RespiratorioDocumento15 pagineAparato RespiratorioDenis ZuloagaNessuna valutazione finora
- Sistema Respiratorio GrupalDocumento5 pagineSistema Respiratorio GrupalisisNessuna valutazione finora
- Sistema Respiratorio HumanoDocumento10 pagineSistema Respiratorio HumanoFrancisco DavalosNessuna valutazione finora
- Biologia Grado Sexto #6Documento5 pagineBiologia Grado Sexto #6Carolina SánchezNessuna valutazione finora
- Tema 4Documento6 pagineTema 4mercedes martinez de toroNessuna valutazione finora
- Sistema RespiratorioDocumento4 pagineSistema RespiratorioJOSUE DAMIAN GONZALEZ SANTAMARIANessuna valutazione finora
- Aparato Respiratorio Experimemnto de CTADocumento75 pagineAparato Respiratorio Experimemnto de CTAJhimy PrincipeNessuna valutazione finora
- Farmacología RespiratoriaDocumento24 pagineFarmacología RespiratoriaRoger MonforteNessuna valutazione finora
- FISIOLOGIA RESPIRATORIA HUMANADocumento27 pagineFISIOLOGIA RESPIRATORIA HUMANAyuglennys amayaNessuna valutazione finora
- Aparato RespiratorioDocumento10 pagineAparato RespiratorioRivera GabrielaNessuna valutazione finora
- Sist. Resp.Documento20 pagineSist. Resp.Surkely GarciaNessuna valutazione finora
- Aparato RespiratorioDocumento18 pagineAparato RespiratorioMaryAngelNessuna valutazione finora
- Aparato RespiratorioDocumento7 pagineAparato RespiratorioAyde Carmita Herrera ArmijosNessuna valutazione finora
- Aparato RespiratorioDocumento8 pagineAparato RespiratorioLuis ChabrillonNessuna valutazione finora
- Introducción al sistema respiratorio humano: estructura, funciones y procesosDocumento8 pagineIntroducción al sistema respiratorio humano: estructura, funciones y procesosKaryJung100% (2)
- Respiración - Wikipedia, La Enciclopedia LibreDocumento28 pagineRespiración - Wikipedia, La Enciclopedia LibreEyilda QuinteroNessuna valutazione finora
- Sistemas Del Cuerpo HumanoDocumento18 pagineSistemas Del Cuerpo HumanoanahimunizaNessuna valutazione finora
- Informe Del Pulmon - WorDocumento18 pagineInforme Del Pulmon - WorDiego EduardoNessuna valutazione finora
- Sistema respiratorioDocumento4 pagineSistema respiratorioSoy UvaNessuna valutazione finora
- Sistema Respiratorio AliDocumento5 pagineSistema Respiratorio AliBrenda SottileNessuna valutazione finora
- Fisiologia Pulmonar (1) (3673)Documento31 pagineFisiologia Pulmonar (1) (3673)Rodolfo Mas TunNessuna valutazione finora
- Sistema RespiratorioDocumento3 pagineSistema RespiratorioMaguiber LozanoNessuna valutazione finora
- La Respiración Permite El Intercambio de Gases Entre El Organismo y El AmbienteDocumento9 pagineLa Respiración Permite El Intercambio de Gases Entre El Organismo y El AmbienteAmy MossNessuna valutazione finora
- Anatomia - FisiologiaDocumento6 pagineAnatomia - FisiologiaEdward AndradeNessuna valutazione finora
- Sistema respiratorio: estructuras y funciones claveDocumento9 pagineSistema respiratorio: estructuras y funciones clavepamela valenzuelaNessuna valutazione finora
- Respiracion 1Documento13 pagineRespiracion 1val ceron samaNessuna valutazione finora
- Istema Espiratorio: Modul ODocumento13 pagineIstema Espiratorio: Modul Ochristian LopezNessuna valutazione finora
- Anatomía R.A 2.1Documento10 pagineAnatomía R.A 2.1Lizbeth GarciaNessuna valutazione finora
- Anatomia y Fisiologia Del Aparato RespiratorioDocumento28 pagineAnatomia y Fisiologia Del Aparato RespiratorioEliezer RiveroNessuna valutazione finora
- Respirator I oDocumento6 pagineRespirator I oAntonia SepulvedaNessuna valutazione finora
- Sistema respiratorio y renal: funciones y partesDocumento17 pagineSistema respiratorio y renal: funciones y partesCatalina MoraNessuna valutazione finora
- Conceptos Y Estrategias En Terapia RespiratoriaDa EverandConceptos Y Estrategias En Terapia RespiratoriaValutazione: 4 su 5 stelle4/5 (1)
- Hemorragia Digestiva PediatriaDocumento42 pagineHemorragia Digestiva PediatriaJesús MejiasNessuna valutazione finora
- Hiperplasia EndometrialDocumento3 pagineHiperplasia EndometrialJesús MejiasNessuna valutazione finora
- Antropometria Pediatrica GeneralidadesDocumento6 pagineAntropometria Pediatrica GeneralidadesJesús MejiasNessuna valutazione finora
- Impacto AmbientalDocumento19 pagineImpacto AmbientalJesús MejiasNessuna valutazione finora
- Distancia de CurvasDocumento1 paginaDistancia de CurvasJesús MejiasNessuna valutazione finora
- Asistencia Médica Durante La Guerra de IndependenciaDocumento13 pagineAsistencia Médica Durante La Guerra de IndependenciaJesús MejiasNessuna valutazione finora
- Contaminacion AmbientalDocumento19 pagineContaminacion AmbientalJesús MejiasNessuna valutazione finora
- Contaminacion AmbientalDocumento19 pagineContaminacion AmbientalJesús MejiasNessuna valutazione finora
- Contam I NacionDocumento17 pagineContam I NacionJesús MejiasNessuna valutazione finora
- Contaminacion AmbientalDocumento19 pagineContaminacion AmbientalJesús MejiasNessuna valutazione finora
- Matriz MefiDocumento5 pagineMatriz MefiVeronica OrtegaNessuna valutazione finora
- Proceso SDocumento10 pagineProceso SMariano Emmanuel León AcostaNessuna valutazione finora
- Plagas de Esparrago Asc PDFDocumento65 paginePlagas de Esparrago Asc PDFJosé LuisNessuna valutazione finora
- Estudio Caso 2Documento4 pagineEstudio Caso 2EDWIN MARTINEZNessuna valutazione finora
- Cronicas de Una CaidaDocumento3 pagineCronicas de Una CaidaTierra RojaNessuna valutazione finora
- Práctica Transferencia de Calor y CalorimetriaDocumento8 paginePráctica Transferencia de Calor y CalorimetriaLuis HernandezNessuna valutazione finora
- Guía de Comandos para IADocumento11 pagineGuía de Comandos para IAIGNACIO SARROCA POBLETENessuna valutazione finora
- Fribroma Traumático. Revisión de LiteraturaDocumento7 pagineFribroma Traumático. Revisión de LiteraturaBerenice VazquezNessuna valutazione finora
- DigimonDocumento2 pagineDigimonAnonymous QRwSJmRsEPNessuna valutazione finora
- LeyClimáticaColombiaDocumento4 pagineLeyClimáticaColombiahugo ferney murillo restrepoNessuna valutazione finora
- Los AnimalesDocumento2 pagineLos AnimalestfdyugfvuighNessuna valutazione finora
- Proyecto Interdiciplinar Q2Documento3 pagineProyecto Interdiciplinar Q2Anonymous 47g7jJQ5Nessuna valutazione finora
- Comparativo Instituciones Nacionales de La Evaluación y Certificación de Calidad Educativa.Documento12 pagineComparativo Instituciones Nacionales de La Evaluación y Certificación de Calidad Educativa.Gaby VBNessuna valutazione finora
- Semaforos Inteligentes v5Documento19 pagineSemaforos Inteligentes v5Cesar Aron Martinez AnayaNessuna valutazione finora
- DO Evaluación y TendenciasDocumento21 pagineDO Evaluación y TendenciasMarie ClaireNessuna valutazione finora
- Hongos y Levaduras1Documento42 pagineHongos y Levaduras1javiera gutierrez leffortNessuna valutazione finora
- 4.procesadores CISC y Procesadores RISCDocumento4 pagine4.procesadores CISC y Procesadores RISCMartin TetlalmatziNessuna valutazione finora
- Cuadernillo DISCALCULIADocumento41 pagineCuadernillo DISCALCULIACarolina Garcia100% (10)
- Manual de Calidad 2 PDFDocumento81 pagineManual de Calidad 2 PDFAvril BurciagaNessuna valutazione finora
- Resumen ID DupontDocumento5 pagineResumen ID DupontHector Espinoza MelgarNessuna valutazione finora
- Evaluacion WISC RDocumento29 pagineEvaluacion WISC RuriceciliaNessuna valutazione finora
- CV Modelo UTPDocumento2 pagineCV Modelo UTPCarlos Navarro100% (1)
- Silabo de Codigo CivilDocumento6 pagineSilabo de Codigo CivilSegundo Ulises Calderón ChambiNessuna valutazione finora
- Estilos de La NegociaciónDocumento11 pagineEstilos de La NegociaciónArleth Thais Rivera VargasNessuna valutazione finora
- Trabajo Práctico PsicoDocumento3 pagineTrabajo Práctico PsicoMauro PascaleNessuna valutazione finora
- Caso 1 - Leo Ticona CanchaDocumento2 pagineCaso 1 - Leo Ticona CanchaLeo André Ticona CanchaNessuna valutazione finora
- Contabilidad FinancieraDocumento103 pagineContabilidad FinancieraEduardo Campos100% (1)
- Guia N°4Taller Números Complejos Definitiva - Terceros A, B y BicentenarioDocumento6 pagineGuia N°4Taller Números Complejos Definitiva - Terceros A, B y BicentenarioAngelo Huaiquil aravenaNessuna valutazione finora
- Examen Enseñanza Por Competencia Semana 3Documento3 pagineExamen Enseñanza Por Competencia Semana 3Jose adrian Rocha rocha33% (3)
- Microeconomia 2, Modelo de PreciosDocumento7 pagineMicroeconomia 2, Modelo de PreciosFrancisco Elissalde MezaNessuna valutazione finora