Potrebbero piacerti anche
- El Tema Del Hombre en Los Escritos de E. G. de White-Shibata VilogrónDocumento124 pagineEl Tema Del Hombre en Los Escritos de E. G. de White-Shibata VilogrónLuis Alberto Marín AguilarNessuna valutazione finora
- Revista Estudios Canarios, 56 (2012)Documento296 pagineRevista Estudios Canarios, 56 (2012)Luis Alberto Marín AguilarNessuna valutazione finora
- Ingenieria en MexicoDocumento401 pagineIngenieria en MexicoPatricia Mc CartneyNessuna valutazione finora
- Teoría y Análisis de La Narrativa en Literatura y en CineDocumento340 pagineTeoría y Análisis de La Narrativa en Literatura y en Cinelcongo100% (4)
- Vladímir Soloviev y La Filosofía-Fernández Calzada-TesisDocumento420 pagineVladímir Soloviev y La Filosofía-Fernández Calzada-TesisLuis Alberto Marín Aguilar100% (2)
- Biografía de Garcia Icazbalceta. Galindo y VillaDocumento46 pagineBiografía de Garcia Icazbalceta. Galindo y VillaLuis Alberto Marín AguilarNessuna valutazione finora
- Para Entender La Militarización en MéxicoDocumento42 paginePara Entender La Militarización en MéxicoLuis Alberto Marín AguilarNessuna valutazione finora
- 05-Tratamiento Manual Del Vertigo PosicionalDocumento10 pagine05-Tratamiento Manual Del Vertigo PosicionalAngie VarasNessuna valutazione finora
- Vestigios Del Cuento Maravilloso. F. MaturanaDocumento10 pagineVestigios Del Cuento Maravilloso. F. MaturanaLuis Alberto Marín AguilarNessuna valutazione finora
- Cuentos de Max AubDocumento426 pagineCuentos de Max AubJuan Carlos100% (1)
- Metodo (Maan Zopa)Documento44 pagineMetodo (Maan Zopa)Claudia Varon100% (1)
- Literatura ArgentinaDocumento45 pagineLiteratura ArgentinaLuis Alberto Marín AguilarNessuna valutazione finora
- Observatorio 13-Seguridad y Violencia en CentroaméricaDocumento234 pagineObservatorio 13-Seguridad y Violencia en CentroaméricaLuis Alberto Marín AguilarNessuna valutazione finora
- Violencia Politica en El Perú 80-88 IDocumento550 pagineViolencia Politica en El Perú 80-88 IDante Bobadilla Ramírez100% (2)
- Serie III - Historia MedievalDocumento581 pagineSerie III - Historia MedievalLuis Alberto Marín Aguilar100% (2)
- Romanticiasmo y MitoDocumento11 pagineRomanticiasmo y MitoBetina KeizmanNessuna valutazione finora
- Ceci y El Mar (Con Posdata)Documento3 pagineCeci y El Mar (Con Posdata)Luis Alberto Marín AguilarNessuna valutazione finora
- Revista Biblioteca de México 114. Alfonso ReyesDocumento68 pagineRevista Biblioteca de México 114. Alfonso ReyesLuis Alberto Marín AguilarNessuna valutazione finora
- Glanz - Astucia Como Novela NacionalDocumento12 pagineGlanz - Astucia Como Novela NacionalJimeno OrozcoNessuna valutazione finora
- Los Puntos Sobre LasiesDocumento189 pagineLos Puntos Sobre LasiesNes Blum100% (1)
- Revista Rocinante 11.Documento88 pagineRevista Rocinante 11.Luis Alberto Marín AguilarNessuna valutazione finora
- Curso de Teoria Politica Eduardo Jorge ArnolettoDocumento393 pagineCurso de Teoria Politica Eduardo Jorge Arnoletto3elizaNessuna valutazione finora
- La Lápida de La Calzada. Graulich-OchoaDocumento26 pagineLa Lápida de La Calzada. Graulich-OchoaLuis Alberto Marín AguilarNessuna valutazione finora
- Flora Vascular de Andalucía Oriental - Tomo - 1. Junta de AndalucíaDocumento436 pagineFlora Vascular de Andalucía Oriental - Tomo - 1. Junta de AndalucíaLuis Alberto Marín AguilarNessuna valutazione finora
- Una Biblioteca Digital de Prensa Antigua. El Caso de Sevilla Ss. XVI-XVIIIDocumento293 pagineUna Biblioteca Digital de Prensa Antigua. El Caso de Sevilla Ss. XVI-XVIIILuis Alberto Marín AguilarNessuna valutazione finora
- Co Solucion EspDocumento75 pagineCo Solucion Esperhetz6686Nessuna valutazione finora
- Biblioteca de México 94. RevistaDocumento68 pagineBiblioteca de México 94. RevistaLuis Alberto Marín AguilarNessuna valutazione finora
- 76 77 1 PBDocumento11 pagine76 77 1 PBaaooggNessuna valutazione finora
- Bioetica.1gDocumento5 pagineBioetica.1gDario IllanesNessuna valutazione finora
- Fisiologia Suprarrenal I Set 2008Documento65 pagineFisiologia Suprarrenal I Set 2008DELIANessuna valutazione finora
- Ranula PDFDocumento5 pagineRanula PDFMich ReynosoNessuna valutazione finora
- CADENA PRODUCTIVA DEL ARROZ - Docx1329559868Documento8 pagineCADENA PRODUCTIVA DEL ARROZ - Docx1329559868Jhon GualNessuna valutazione finora
- 7ma Clase - Quinolonas, Sulfonamidas, Trimetoprim. Dra. de FreitasDocumento39 pagine7ma Clase - Quinolonas, Sulfonamidas, Trimetoprim. Dra. de FreitasCátedra de Farmacología de la Escuela de Medicina de La UNIVERSIDAD DEL ZULIA100% (2)
- Clase de Biologia 02.07.20Documento15 pagineClase de Biologia 02.07.20INDUSTRIAL SERVICESNessuna valutazione finora
- Manual CTI8 - 11Documento24 pagineManual CTI8 - 11Marco Corriols Caldera100% (1)
- Toxicología GeneralidadesDocumento31 pagineToxicología GeneralidadesEmilio GarciaNessuna valutazione finora
- Caso Practico Comunicacion EmpresarialDocumento7 pagineCaso Practico Comunicacion EmpresarialJennifer Yepes BrunNessuna valutazione finora
- DaltonismoDocumento3 pagineDaltonismosolcito_ceballos100% (1)
- Clasificación TaxonómicaDocumento8 pagineClasificación TaxonómicaFelipe B MendozaNessuna valutazione finora
- Bebidas EnergéticasDocumento7 pagineBebidas EnergéticasDiego Torres100% (1)
- Matriz Iper RestaurantDocumento6 pagineMatriz Iper Restaurantlaboratorios0% (1)
- Diarrea Cronica ExposicionDocumento23 pagineDiarrea Cronica ExposicionRosa Laura MacíasNessuna valutazione finora
- 6 Tesis Pie Diabético Ultima OkDocumento32 pagine6 Tesis Pie Diabético Ultima OksimonNessuna valutazione finora
- Pretest Rm3v Salud Publica Sin ClaveDocumento3 paginePretest Rm3v Salud Publica Sin ClavesaidajaraNessuna valutazione finora
- Tema 7 - Ventilación Mecánica No Invasiva en El Covid 19 Cuándo Utilizarlos - Cómo Utilizarlos - Dónde UtilizarlosDocumento18 pagineTema 7 - Ventilación Mecánica No Invasiva en El Covid 19 Cuándo Utilizarlos - Cómo Utilizarlos - Dónde UtilizarlosWilson Torres HuNessuna valutazione finora
- Triptico para Un Regreso A Clases SeguroDocumento2 pagineTriptico para Un Regreso A Clases Seguroabarbaralm85% (13)
- Plantas Ancestrales y Medicinales Clase # 11aDocumento9 paginePlantas Ancestrales y Medicinales Clase # 11aJunior FigueroaNessuna valutazione finora
- PEDIATRIADocumento19 paginePEDIATRIAWaldir Urbina EscobarNessuna valutazione finora
- Ficha Clinica ImprimirrrDocumento7 pagineFicha Clinica ImprimirrrSYLVER CH QQNessuna valutazione finora
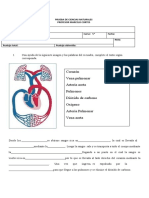
- Prueba 5° Ciencias CasaDocumento2 paginePrueba 5° Ciencias CasaMarcelo Cortes Brito100% (1)
- 8 Capitulo Portada 75Documento3 pagine8 Capitulo Portada 75Roberto Rouhana MonagasNessuna valutazione finora
- P Engormax PDFDocumento1 paginaP Engormax PDFEdhisson Javier100% (1)
- La ChusDocumento1 paginaLa ChusMario Garcia-riscoNessuna valutazione finora
- Proyecto Final Gestión en Prevención de Riesgos IaccDocumento6 pagineProyecto Final Gestión en Prevención de Riesgos IaccCristhianHananíasEventos60% (5)
- Tarea 1 de Psicopatologia 2Documento7 pagineTarea 1 de Psicopatologia 2miguelNessuna valutazione finora
- Curriculum 2011 FabianDocumento5 pagineCurriculum 2011 FabianJorge Blanco BaldiviesoNessuna valutazione finora
- Vive 100Documento2 pagineVive 100Diana Laura100% (1)
- ESPOROTRICOSIS Caso ClinicoDocumento35 pagineESPOROTRICOSIS Caso ClinicoBreilin PosadasNessuna valutazione finora
- Los Secretos De La Mente Millonaria: Domina el juego de la riquezaDa EverandLos Secretos De La Mente Millonaria: Domina el juego de la riquezaValutazione: 5 su 5 stelle5/5 (457)
- Recupera tu mente, reconquista tu vidaDa EverandRecupera tu mente, reconquista tu vidaValutazione: 5 su 5 stelle5/5 (7)
- Psicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaDa EverandPsicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaValutazione: 4.5 su 5 stelle4.5/5 (766)
- El Monje Que Vendio Su Ferrari: Una Fábula EspiritualDa EverandEl Monje Que Vendio Su Ferrari: Una Fábula EspiritualValutazione: 4.5 su 5 stelle4.5/5 (1699)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaDa EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaValutazione: 5 su 5 stelle5/5 (1872)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadDa EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadValutazione: 4.5 su 5 stelle4.5/5 (1831)
- Signos y símbolos: Desvelando el significado espiritual de los sellos angélicos, tótems y otros símbolos mágicos, sagrados y religiososDa EverandSignos y símbolos: Desvelando el significado espiritual de los sellos angélicos, tótems y otros símbolos mágicos, sagrados y religiososValutazione: 5 su 5 stelle5/5 (1)
- Yo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Da EverandYo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Valutazione: 5 su 5 stelle5/5 (7)
- Resetea tu mente. Descubre de lo que eres capazDa EverandResetea tu mente. Descubre de lo que eres capazValutazione: 5 su 5 stelle5/5 (196)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDa EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaValutazione: 5 su 5 stelle5/5 (203)
- El poder del optimismo: Herramientas para vivir de forma más positivaDa EverandEl poder del optimismo: Herramientas para vivir de forma más positivaValutazione: 5 su 5 stelle5/5 (16)
- ¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadDa Everand¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadValutazione: 5 su 5 stelle5/5 (198)
- Fuerza de voluntad: Cómo practicar y dominar el autocontrolDa EverandFuerza de voluntad: Cómo practicar y dominar el autocontrolValutazione: 4.5 su 5 stelle4.5/5 (90)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDa EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanValutazione: 4.5 su 5 stelle4.5/5 (64)
- Guías Espirituales: ¡Una guía para conectarte y comunicarte con tus guías espirituales!Da EverandGuías Espirituales: ¡Una guía para conectarte y comunicarte con tus guías espirituales!Valutazione: 4.5 su 5 stelle4.5/5 (14)
- Influencia. La psicología de la persuasiónDa EverandInfluencia. La psicología de la persuasiónValutazione: 4.5 su 5 stelle4.5/5 (14)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDa EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceValutazione: 5 su 5 stelle5/5 (8)
- Entrenamiento Científico con pesas: Fitness InteligenteDa EverandEntrenamiento Científico con pesas: Fitness InteligenteValutazione: 5 su 5 stelle5/5 (3)
- En sintonía con tu ciclo femenino: FLO. Aprende a sincronizarte con tu bioquímica para dar rienda suelta a tu creatividad, mejorar tu vida y hacer más con menos estrésDa EverandEn sintonía con tu ciclo femenino: FLO. Aprende a sincronizarte con tu bioquímica para dar rienda suelta a tu creatividad, mejorar tu vida y hacer más con menos estrésValutazione: 4.5 su 5 stelle4.5/5 (11)
- Homo antecessor: El nacimiento de una especieDa EverandHomo antecessor: El nacimiento de una especieValutazione: 5 su 5 stelle5/5 (1)
- ¡Basta ya de ser un Tipo Lindo! (No More Mr. Nice Guy): Un Plan Probado para Que Obtengas Lo Que Quieras en La Vida El Sexo y El Amor (A Proven Plan for Getting What You Want in Love, Sex and Life)Da Everand¡Basta ya de ser un Tipo Lindo! (No More Mr. Nice Guy): Un Plan Probado para Que Obtengas Lo Que Quieras en La Vida El Sexo y El Amor (A Proven Plan for Getting What You Want in Love, Sex and Life)Valutazione: 5 su 5 stelle5/5 (47)
- Tu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaDa EverandTu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaValutazione: 5 su 5 stelle5/5 (2)
- No desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaDa EverandNo desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaValutazione: 5 su 5 stelle5/5 (13)