Potrebbero piacerti anche
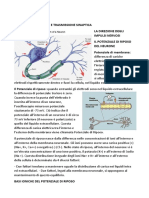
- 4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azioneDocumento43 pagine4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azionedavide di lorenzoNessuna valutazione finora
- L'impulso NervosoDocumento7 pagineL'impulso NervosoGiancarlo AttivissimoNessuna valutazione finora
- 10 - Potenziali Graduati e Potenziale D-AzioneDocumento48 pagine10 - Potenziali Graduati e Potenziale D-AzioneMarcoNessuna valutazione finora
- 2 FondamentiDocumento42 pagine2 FondamentiAndrewNessuna valutazione finora
- Conduzione Impulso NervosoDocumento32 pagineConduzione Impulso NervosoMarcoNessuna valutazione finora
- Trasmissione Sinaptica e Integrazione NeuronaleDocumento11 pagineTrasmissione Sinaptica e Integrazione NeuronaleAnnamaria MaderaNessuna valutazione finora
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- Cap 4 Riassunto FinDocumento13 pagineCap 4 Riassunto Finandrorru.stefanoNessuna valutazione finora
- I Recettori SensorialiDocumento37 pagineI Recettori SensorialiMaryna100% (1)
- Lezione 2 fisiologiaDocumento47 pagineLezione 2 fisiologiamarticatalin8Nessuna valutazione finora
- Psico CognitivaDocumento41 paginePsico CognitivaMichele CarontiNessuna valutazione finora
- Appunti Di PsicobiologiaDocumento21 pagineAppunti Di Psicobiologialucatinaro1997Nessuna valutazione finora
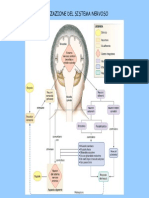
- S NervosoDocumento104 pagineS NervosoAgostino InzerilloNessuna valutazione finora
- Effetti Fisiologici Derivanti Dall'Assunzione Di MDMADocumento31 pagineEffetti Fisiologici Derivanti Dall'Assunzione Di MDMAhellenNessuna valutazione finora
- Lezioni Di NeurologiaDocumento122 pagineLezioni Di NeurologiaTania NocchieroNessuna valutazione finora
- Cap 5 Riassunto FinDocumento11 pagineCap 5 Riassunto Finandrorru.stefanoNessuna valutazione finora
- 1 Codifica Delle Informazioni SensorialiDocumento6 pagine1 Codifica Delle Informazioni SensorialiMarco Bronzini100% (1)
- RetinaDocumento34 pagineRetinaapi-3704348100% (4)
- La Plasticità SinapsiDocumento46 pagineLa Plasticità SinapsiGabriele GargiuloNessuna valutazione finora
- Fisiologia Lezione 6Documento2 pagineFisiologia Lezione 6The last chimeraNessuna valutazione finora
- Capitolo 2 Purves NeuroscienzeDocumento4 pagineCapitolo 2 Purves NeuroscienzeElena MaestroniNessuna valutazione finora
- Sistema NervosoDocumento23 pagineSistema NervosoDiana ChiccaNessuna valutazione finora
- 1.sistemi Sensoriali 01Documento23 pagine1.sistemi Sensoriali 01larisutzafNessuna valutazione finora
- 5 Potenziali ElettrotoniciDocumento8 pagine5 Potenziali ElettrotoniciClaudia OrlannoNessuna valutazione finora
- Mod Appunti Fondamenti Di FisiologiaDocumento7 pagineMod Appunti Fondamenti Di Fisiologiaandreadiamante85Nessuna valutazione finora
- Conduzione NervosaDocumento10 pagineConduzione NervosaMiriam LongobardiNessuna valutazione finora
- Fisiologia AnimaleDocumento11 pagineFisiologia AnimaleGIULIA MICALETTONessuna valutazione finora
- Discriminazione Tra Due PuntiDocumento25 pagineDiscriminazione Tra Due PuntiFederica ManziNessuna valutazione finora
- Domande PIT FisiologiaDocumento5 pagineDomande PIT FisiologiaJoshuaKKyleNessuna valutazione finora
- 1 Potenziale Di Equilibrio Di NernstDocumento5 pagine1 Potenziale Di Equilibrio Di NernstClaudia OrlannoNessuna valutazione finora
- Marieb Sintesi 62091 c14Documento3 pagineMarieb Sintesi 62091 c14Caterina NuzzoNessuna valutazione finora
- Anestetici LocaliDocumento10 pagineAnestetici LocaliChiaraPetraroliNessuna valutazione finora
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- Il Sistema NervosoDocumento6 pagineIl Sistema NervosoMartina MusmeciNessuna valutazione finora
- Appunti Modulo 1-2-3Documento84 pagineAppunti Modulo 1-2-3ERIKA BERNOCCHINessuna valutazione finora
- 2.impulso NervosoDocumento10 pagine2.impulso NervosofNessuna valutazione finora
- Domande e Risposte Sistema NervosoDocumento4 pagineDomande e Risposte Sistema Nervosoedup220% (1)
- (Medicina-Ita) Neurologia - EEGDocumento44 pagine(Medicina-Ita) Neurologia - EEGedosuper1Nessuna valutazione finora
- Fisiologia Lezione 5Documento3 pagineFisiologia Lezione 5The last chimeraNessuna valutazione finora
- Tutti UnitiDocumento120 pagineTutti UnitiERIKA BERNOCCHINessuna valutazione finora
- Lezione-4-fisiologiaDocumento40 pagineLezione-4-fisiologiamarticatalin8Nessuna valutazione finora
- Olom-OraDocumento8 pagineOlom-OrariccardosalviNessuna valutazione finora
- 3-Udito e PsicoacusticaDocumento9 pagine3-Udito e PsicoacusticaFedericoNessuna valutazione finora
- Psicologia Dell AttenzioneDocumento101 paginePsicologia Dell AttenzioneMarco MenguzzatoNessuna valutazione finora
- Cronopsicologia SbobinatureDocumento182 pagineCronopsicologia Sbobinaturemissright98Nessuna valutazione finora
- Anestesia LocoregionaleDocumento36 pagineAnestesia LocoregionaleLucatntNessuna valutazione finora
- Libro I Confini Di BabeleDocumento10 pagineLibro I Confini Di BabeleLuthienNessuna valutazione finora
- Tessuto NervosoDocumento7 pagineTessuto NervosoeristelaNessuna valutazione finora
- Neurologia SicilianoDocumento247 pagineNeurologia SicilianofilippobNessuna valutazione finora
- Lezione AnestesiaDocumento117 pagineLezione AnestesiareeredNessuna valutazione finora
- BiologiaDocumento6 pagineBiologiaracheleNessuna valutazione finora
- Potenziale D'azioneDocumento30 paginePotenziale D'azioneRiccardo Mason TroiaNessuna valutazione finora
- Fisiologia Cellulare Potenziale AzioneDocumento31 pagineFisiologia Cellulare Potenziale Azionesimo_pNessuna valutazione finora
- Fisiologia Dell' Esercizio e Dello SportDocumento65 pagineFisiologia Dell' Esercizio e Dello SportDaniele Nervi33% (3)
- Fisiologia ValenteDocumento4 pagineFisiologia ValenteFrancesco FalconieriNessuna valutazione finora
- Teoria SinapticaDocumento5 pagineTeoria SinapticaFran BisoNessuna valutazione finora
- Fisiologia - Fisica Dell'EcgDocumento23 pagineFisiologia - Fisica Dell'EcgBenjaminMeghnagi0% (1)
- TESINA - Ins. Fisica Tecnica e Impianti Tecnici - RenzulloDocumento11 pagineTESINA - Ins. Fisica Tecnica e Impianti Tecnici - RenzulloAndrea RenzulloNessuna valutazione finora
- Le correnti a radiofrequenza applicate alla terapia del doloreDa EverandLe correnti a radiofrequenza applicate alla terapia del doloreNessuna valutazione finora
- Medicina 2003-04Documento5 pagineMedicina 2003-04OrlandoCialliNessuna valutazione finora
- Medicina 2000-01Documento3 pagineMedicina 2000-01OrlandoCialliNessuna valutazione finora
- Medicina 1999-00Documento15 pagineMedicina 1999-00OrlandoCialliNessuna valutazione finora
- Simulazione CHIMICA ORGANICADocumento6 pagineSimulazione CHIMICA ORGANICAOrlandoCialliNessuna valutazione finora
- Medicina 1998-99Documento9 pagineMedicina 1998-99OrlandoCialliNessuna valutazione finora
- Medicina 2001-02Documento5 pagineMedicina 2001-02OrlandoCialliNessuna valutazione finora
- Medicina 1997-98Documento9 pagineMedicina 1997-98OrlandoCialliNessuna valutazione finora
- La Via Degli Dei PDFDocumento36 pagineLa Via Degli Dei PDFOrlandoCialli50% (2)
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- TICKET GL200362483sDocumento11 pagineTICKET GL200362483sOrlandoCialliNessuna valutazione finora
- Simulazione CHIMICA ORGANICADocumento6 pagineSimulazione CHIMICA ORGANICAOrlandoCialliNessuna valutazione finora
- Fisiologia Generale 7 PDFDocumento16 pagineFisiologia Generale 7 PDFOrlandoCialliNessuna valutazione finora
- ORLANDO CIALLI 1362520296sDocumento1 paginaORLANDO CIALLI 1362520296sOrlandoCialliNessuna valutazione finora
- Clara Bartalesi 892020472Documento2 pagineClara Bartalesi 892020472OrlandoCialliNessuna valutazione finora
- 2) La Cellula VegetaleDocumento39 pagine2) La Cellula VegetaleOrlandoCialliNessuna valutazione finora
- 2modulo Autodichiarazione 17 3 2020 PDFDocumento1 pagina2modulo Autodichiarazione 17 3 2020 PDFOrlandoCialliNessuna valutazione finora
- Fisiologia Generale 2Documento26 pagineFisiologia Generale 2OrlandoCialliNessuna valutazione finora
- Fisiologia Generale 3Documento31 pagineFisiologia Generale 3OrlandoCialliNessuna valutazione finora
- 4 La Reologia: 4.1 IntroduzioneDocumento26 pagine4 La Reologia: 4.1 IntroduzioneOrlandoCialliNessuna valutazione finora
- Fisiologia Generale 2Documento26 pagineFisiologia Generale 2OrlandoCialliNessuna valutazione finora
- Fisiologia Generale 1Documento41 pagineFisiologia Generale 1OrlandoCialliNessuna valutazione finora
- IR ATR Lab 222 20190910: Liofilizzato N 60°CDocumento4 pagineIR ATR Lab 222 20190910: Liofilizzato N 60°COrlandoCialliNessuna valutazione finora
- RISULTATI ListaDocumento3 pagineRISULTATI ListaOrlandoCialliNessuna valutazione finora
- Es ChimDocumento1 paginaEs ChimOrlandoCialliNessuna valutazione finora
- FiatiDocumento2 pagineFiatiOrlandoCialliNessuna valutazione finora
- RISULTATI ListaDocumento3 pagineRISULTATI ListaOrlandoCialliNessuna valutazione finora
- OrganicaDocumento2 pagineOrganicaOrlandoCialliNessuna valutazione finora
- Resoconto IR e NMRDocumento16 pagineResoconto IR e NMROrlandoCialliNessuna valutazione finora
- Lavoro Steudel AcDocumento4 pagineLavoro Steudel AcOrlandoCialliNessuna valutazione finora
- Approccio Terapeutico RiabilitativoDocumento46 pagineApproccio Terapeutico RiabilitativoBasilio AroneNessuna valutazione finora
- Domande Esame FisiologiaDocumento4 pagineDomande Esame Fisiologiatonino sperdutiNessuna valutazione finora
- Curarsi Con Il MagnesioDocumento13 pagineCurarsi Con Il MagnesiovittaniNessuna valutazione finora
- CALAMAI Scienza Dell'AlimentazioneDocumento56 pagineCALAMAI Scienza Dell'AlimentazioneladydaladyNessuna valutazione finora
- ALLENAMENTO A CASA Creare Il Tuo Fisico PerfettoDocumento85 pagineALLENAMENTO A CASA Creare Il Tuo Fisico PerfettoWaltNessuna valutazione finora
- 8.valutazione Funzionalita' CardiovascolareDocumento8 pagine8.valutazione Funzionalita' Cardiovascolaremelissa calabreseNessuna valutazione finora
- StregoneDocumento20 pagineStregoneStefania BorelloNessuna valutazione finora
- APPUNTI Di Biomeccanica AA 2011 - 12Documento142 pagineAPPUNTI Di Biomeccanica AA 2011 - 12Gerlando Russo IntroitoNessuna valutazione finora
- Miofasciale PDFDocumento35 pagineMiofasciale PDFcarne23Nessuna valutazione finora
- La Riabilitazione Dell - Articolazione Temporo Mandibolare Con Terapia ManualeDocumento58 pagineLa Riabilitazione Dell - Articolazione Temporo Mandibolare Con Terapia ManualeHermann100% (2)
- ORTOPEDIADocumento106 pagineORTOPEDIATiziano LicataNessuna valutazione finora
- Fisiologia Del MuscoloDocumento44 pagineFisiologia Del MuscoloRiccardoMazzaNessuna valutazione finora
- Domande Fisiologia 2 2008-2012Documento16 pagineDomande Fisiologia 2 2008-2012m.pisanoNessuna valutazione finora
- 05 Gnatiferi - Nemertini - CamptozoiDocumento13 pagine05 Gnatiferi - Nemertini - CamptozoiEugenio StracuzzaNessuna valutazione finora
- Dismetrie Arti InferioriDocumento7 pagineDismetrie Arti InferioriGiovanni Russo100% (1)
- Natural Body BuildingDocumento67 pagineNatural Body BuildingMoretti AntoninoNessuna valutazione finora
- Sistema Vestibolare, Funzioni Superiori Dell'encefalo, LinguaggioDocumento17 pagineSistema Vestibolare, Funzioni Superiori Dell'encefalo, Linguaggioapi-3704348Nessuna valutazione finora
- 165 Consigli Di David Russell PDFDocumento24 pagine165 Consigli Di David Russell PDFAlbertoFalcioneNessuna valutazione finora
- Esercizi Per Gli OcchiDocumento4 pagineEsercizi Per Gli OcchiMario LombreschiNessuna valutazione finora
- Differenziamento Cellulare - Derivazione Embrionale IstologiaDocumento17 pagineDifferenziamento Cellulare - Derivazione Embrionale IstologiaWicked_KnightNessuna valutazione finora