Potrebbero piacerti anche
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- FIFA Soccer 11 Plus ManualDocumento76 pagineFIFA Soccer 11 Plus ManualtheroguetrainerNessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Miller Big Blue 400d and 500x Deutz Part ADocumento98 pagineMiller Big Blue 400d and 500x Deutz Part ALUIS ALFREDO SIERRA JUAREZ0% (1)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- 2011 PJ Medical HandbookDocumento210 pagine2011 PJ Medical Handbookdocschneid100% (9)
- IntroBooklet PDFDocumento19 pagineIntroBooklet PDFDaniel KKKNessuna valutazione finora
- Biomechanics of SpineDocumento41 pagineBiomechanics of SpineMuhammad MustaqeemNessuna valutazione finora
- Legal & Other Requirement Rev 00Documento40 pagineLegal & Other Requirement Rev 00Mohd Zulhaidy100% (1)
- RA (Working at Height)Documento4 pagineRA (Working at Height)Yan Naing HtooNessuna valutazione finora
- Helicopter Fire Fighting and RescueDocumento5 pagineHelicopter Fire Fighting and Rescuermaffireschool100% (1)
- Ethicon Wound Closure ManualDocumento126 pagineEthicon Wound Closure Manualkeish193100% (3)
- Accuvac Rescue 16136 enDocumento52 pagineAccuvac Rescue 16136 ensmurd112Nessuna valutazione finora
- Football Sports ProjectDocumento57 pagineFootball Sports ProjectOm Shinde67% (3)
- Supracondylar Humerus FractureDocumento13 pagineSupracondylar Humerus FracturedhapitstinNessuna valutazione finora
- Injuries To The PeriodontiumDocumento3 pagineInjuries To The PeriodontiumIris BakerNessuna valutazione finora
- GI Anatomy Golden!Documento39 pagineGI Anatomy Golden!Ayodeji SotimehinNessuna valutazione finora
- Allied Signal, Inc., Third-Party v. Acme Service Corp., Third-Party, 946 F.2d 1295, 3rd Cir. (1991)Documento3 pagineAllied Signal, Inc., Third-Party v. Acme Service Corp., Third-Party, 946 F.2d 1295, 3rd Cir. (1991)Scribd Government DocsNessuna valutazione finora
- Vasyl Stefanyk - A STONE CROSSDocumento5 pagineVasyl Stefanyk - A STONE CROSSandreusDADANessuna valutazione finora
- Anatomy of Thoracic WallDocumento27 pagineAnatomy of Thoracic WallSamuel Pola Karta Sembiring100% (1)
- BCSW - S1E9 Support PackDocumento5 pagineBCSW - S1E9 Support PackNat ShattNessuna valutazione finora
- Petitioner vs. vs. Respondents: en BancDocumento6 paginePetitioner vs. vs. Respondents: en BancJessica Magsaysay CrisostomoNessuna valutazione finora
- 2nd Grading Test Science 5Documento7 pagine2nd Grading Test Science 5LeonorBagnisonNessuna valutazione finora
- Exercise of Body MechanicsDocumento4 pagineExercise of Body Mechanicslostloved7Nessuna valutazione finora
- SurgeryDocumento135 pagineSurgeryAndi AlfianNessuna valutazione finora
- Risk of Injury in Side Facing Aircraft Seats 200713 CAMI SFSDocumento32 pagineRisk of Injury in Side Facing Aircraft Seats 200713 CAMI SFStjgNessuna valutazione finora
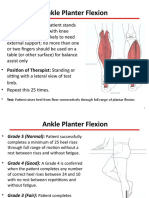
- Ankle Planter Flexion: Test: Patient Raises Heel From Floor Consecutively Through Full Range of Plantar FlexionDocumento30 pagineAnkle Planter Flexion: Test: Patient Raises Heel From Floor Consecutively Through Full Range of Plantar FlexionHasan KhawaldehNessuna valutazione finora
- Waqar Cheema Advocate 5 - 10 - 2019Documento1 paginaWaqar Cheema Advocate 5 - 10 - 2019Akhtar RasoolNessuna valutazione finora
- Worm The Game Edit of Wildbow'sDocumento16 pagineWorm The Game Edit of Wildbow'sLuke AjemNessuna valutazione finora
- H Rac Risk Assessment ActivityDocumento5 pagineH Rac Risk Assessment ActivityroshanrajasNessuna valutazione finora
- Lecture No.2 Axillary Nerve by DR Chaman Lal PTDocumento23 pagineLecture No.2 Axillary Nerve by DR Chaman Lal PTDr Chaman Lal PTNessuna valutazione finora
- First AidDocumento8 pagineFirst AidVikashNessuna valutazione finora
- Commissioner Appointment Petition in Mvop-388-2016-3Documento5 pagineCommissioner Appointment Petition in Mvop-388-2016-3venkatapavankumarNessuna valutazione finora