Potrebbero piacerti anche
- Qué Es El MetabolismoDocumento8 pagineQué Es El MetabolismoJosé BlondellNessuna valutazione finora
- Clasificación Del MetabolismoDocumento7 pagineClasificación Del MetabolismoAna Gabriela Hans SalazarNessuna valutazione finora
- Taller NutriciónDocumento11 pagineTaller NutriciónFerney LópezNessuna valutazione finora
- El MetabolismoDocumento8 pagineEl MetabolismoLaura PerezNessuna valutazione finora
- Metabolismo Anabolismo y CatabolismoDocumento21 pagineMetabolismo Anabolismo y CatabolismoJose Nicolás Valdez BurgosNessuna valutazione finora
- Bme U1 A1 DoccDocumento6 pagineBme U1 A1 DoccDorisliliaNessuna valutazione finora
- METABOLISMODocumento17 pagineMETABOLISMOJENI MARCILLO MORÁNNessuna valutazione finora
- El Metabolismo Del Ser HumanoDocumento25 pagineEl Metabolismo Del Ser HumanoFernando GutierrezNessuna valutazione finora
- Metabolismo y NutricionDocumento10 pagineMetabolismo y Nutricionweli strongerNessuna valutazione finora
- Biologia Grupo 6 MetabolismoDocumento5 pagineBiologia Grupo 6 MetabolismoJHARIM NOELY AUCCAPIÑA GUILLENNessuna valutazione finora
- MetabolismoDocumento12 pagineMetabolismoYisus xdNessuna valutazione finora
- Cortés González Juan Pablo - Taller Celula y Nutricion CORRECCIÓNDocumento8 pagineCortés González Juan Pablo - Taller Celula y Nutricion CORRECCIÓNJuan Pablo CortésNessuna valutazione finora
- Cuadro Comparativo Anabolismo y CatabolismoDocumento3 pagineCuadro Comparativo Anabolismo y CatabolismoKelly Briseño Lopez100% (2)
- MetabolismoDocumento5 pagineMetabolismoAngela Oñate HaroNessuna valutazione finora
- Metabolismo de Los NutrientesDocumento103 pagineMetabolismo de Los NutrientesEvilin Romero Marchan100% (1)
- Metabolismo de BioquimicaDocumento14 pagineMetabolismo de BioquimicaAnthony DiazNessuna valutazione finora
- Do You Consider Yourself Peruvian Yes, No, Why Yes, Because I Was Born in Wonderful Peru and I Also Have Cultures, Religions, I Have Peruvian Blood.Documento2 pagineDo You Consider Yourself Peruvian Yes, No, Why Yes, Because I Was Born in Wonderful Peru and I Also Have Cultures, Religions, I Have Peruvian Blood.Glenda Cordova GarciaNessuna valutazione finora
- METABOLISMODocumento9 pagineMETABOLISMOingpablosolisNessuna valutazione finora
- Tarea 9 Biologia DigestionDocumento3 pagineTarea 9 Biologia DigestionNicoleNessuna valutazione finora
- Metabolismo de Animales Monogástricos y RumiantesDocumento14 pagineMetabolismo de Animales Monogástricos y RumiantesDaniel de Jesús Lopez GutiérrezNessuna valutazione finora
- Procesos Metabólicos de Obtención de EnergíaDocumento20 pagineProcesos Metabólicos de Obtención de EnergíaRocola LocaNessuna valutazione finora
- El MetabolismoDocumento2 pagineEl MetabolismoIsrael AndresNessuna valutazione finora
- Etapas o Fases Del MetabolismoDocumento8 pagineEtapas o Fases Del MetabolismoFlorr VerasayNessuna valutazione finora
- Investigacion de QuimicaDocumento5 pagineInvestigacion de QuimicaMaryNessuna valutazione finora
- LeslyDocumento12 pagineLeslyJoselyn Liza (Peruana en colombia)Nessuna valutazione finora
- SEMANA Nro. 07 METABOLISMO Y RESPIRACIÓN CELULAR METABOLISMODocumento52 pagineSEMANA Nro. 07 METABOLISMO Y RESPIRACIÓN CELULAR METABOLISMOAna Estela Carbonel NeciosupNessuna valutazione finora
- Sara Ciencias InfDocumento19 pagineSara Ciencias InfLaura Dhayana Saray SerratoNessuna valutazione finora
- Metabolismo y FotosíntesisDocumento9 pagineMetabolismo y FotosíntesisJacqueline FranciscoNessuna valutazione finora
- METABOLISMO DE LOS NUTRIENTES Resumen BromatologíaDocumento11 pagineMETABOLISMO DE LOS NUTRIENTES Resumen BromatologíaIván Narváez AguilarNessuna valutazione finora
- MetabolismoDocumento4 pagineMetabolismoRa FaNessuna valutazione finora
- El MetabolismoDocumento3 pagineEl MetabolismoCarla GonzalezNessuna valutazione finora
- Anabolismo y CatabolismoDocumento4 pagineAnabolismo y CatabolismoAna GabrielaNessuna valutazione finora
- Comer Sano para Vivir MejorDocumento53 pagineComer Sano para Vivir MejorNICA FERNANDEZNessuna valutazione finora
- 8 Introducción A MetabolismoDocumento18 pagine8 Introducción A MetabolismoJasna ZovakNessuna valutazione finora
- Conceptos Básicos Sobre El Metabolismo Digestivo - LALBDocumento6 pagineConceptos Básicos Sobre El Metabolismo Digestivo - LALBLibni Aridai LBNessuna valutazione finora
- MetabolismoDocumento2 pagineMetabolismoJairo Jose Menendez RamirezNessuna valutazione finora
- Taller BioquimicaDocumento6 pagineTaller BioquimicaValentina NuñezNessuna valutazione finora
- El Metabolismo-Oriannys Franco-Seccion 18Documento10 pagineEl Metabolismo-Oriannys Franco-Seccion 18Obdulis FrancoNessuna valutazione finora
- MetabolismoDocumento16 pagineMetabolismoPiere Anthony Amaya GonzalesNessuna valutazione finora
- Glosario de Conceptos Básicos de BioquimicaDocumento55 pagineGlosario de Conceptos Básicos de BioquimicaJonathan Rosas Pesina100% (3)
- El MetabolismoDocumento13 pagineEl MetabolismoErnest SantiagoNessuna valutazione finora
- Fico 2Documento63 pagineFico 2JuniorFloresNessuna valutazione finora
- El Proceso de Digestión, Absorción y Metabolización de Nutrientes.Documento19 pagineEl Proceso de Digestión, Absorción y Metabolización de Nutrientes.Alezie Roosevelt Paul GarciaNessuna valutazione finora
- CT - Experiencia VIII Actividad 3Documento4 pagineCT - Experiencia VIII Actividad 3Katia Quispe RamirezNessuna valutazione finora
- Tema #4Documento8 pagineTema #4YULIANA AYMEE PEA RODRIGUEZNessuna valutazione finora
- Metabolismo y NutriciónDocumento21 pagineMetabolismo y NutriciónMore ParedesNessuna valutazione finora
- Biología - MetabolismoDocumento3 pagineBiología - MetabolismoBrianny MoralesNessuna valutazione finora
- METABOLISMODocumento15 pagineMETABOLISMOkdserra27Nessuna valutazione finora
- TALLER - No. - 1 - de - BIOQUIMICA - 2021 - 2 - DEFDocumento28 pagineTALLER - No. - 1 - de - BIOQUIMICA - 2021 - 2 - DEFlaura pardoNessuna valutazione finora
- A.Resumen Principios de BioenergéticaDocumento4 pagineA.Resumen Principios de BioenergéticaAlfonso AlvarezNessuna valutazione finora
- Metabolism oDocumento8 pagineMetabolism omartin lunaNessuna valutazione finora
- Concepto MetabolismoDocumento4 pagineConcepto Metabolismobider andres monroy leytonNessuna valutazione finora
- TRabajo de BiologiaDocumento5 pagineTRabajo de BiologiaCeleste RamosNessuna valutazione finora
- Presentacion pt2Documento38 paginePresentacion pt2Jahir CarrilloNessuna valutazione finora
- 2-Metabolismo Atp...Documento6 pagine2-Metabolismo Atp...Fabio RiverosNessuna valutazione finora
- Investigacion DirigidaDocumento10 pagineInvestigacion DirigidaJana Misseth Gaxiola CastroNessuna valutazione finora
- Metabolismo Minerales y VitaminasDocumento8 pagineMetabolismo Minerales y VitaminasJohana Pineda100% (1)
- Act2 - U3 - Ciencias de La Salud IDocumento12 pagineAct2 - U3 - Ciencias de La Salud IRenata MonroyNessuna valutazione finora
- La Ciencia Del Metabolismo: Metabolismo sin rodeosDa EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosValutazione: 5 su 5 stelle5/5 (1)
- Nutrición: La solución para vivir sin enfermedadesDa EverandNutrición: La solución para vivir sin enfermedadesNessuna valutazione finora
- TintoreriaDocumento11 pagineTintoreriaFlorence VillarroelNessuna valutazione finora
- Tipos de Curado de ConcretoDocumento3 pagineTipos de Curado de ConcretoJames NilNessuna valutazione finora
- FISICADocumento2 pagineFISICAWilcer Cusquisiban De La CruzNessuna valutazione finora
- FundamentosDocumento6 pagineFundamentosJenisser Avalos VasquezNessuna valutazione finora
- Informe SEMDocumento9 pagineInforme SEMFabricio Castillo Avilez0% (1)
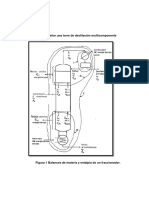
- Pasos para Diseñar Una Torre de Destilación MulticomponenteDocumento9 paginePasos para Diseñar Una Torre de Destilación MulticomponenteGasersithoOnneNessuna valutazione finora
- Descripcion Del Proceso de RonDocumento14 pagineDescripcion Del Proceso de RonRobmar Alvarez50% (6)
- Naturaleza Del Calor y La TemperaturaDocumento9 pagineNaturaleza Del Calor y La TemperaturagustavoNessuna valutazione finora
- Electrometalurgia A. MoralesDocumento93 pagineElectrometalurgia A. Moralescenthus100% (3)
- SolucionesDocumento4 pagineSolucionesbfv100% (1)
- Taller-1-SOLUCIONES-VOLUMETRÍA-GRAVIMETRÍA (Pre-Parcial)Documento3 pagineTaller-1-SOLUCIONES-VOLUMETRÍA-GRAVIMETRÍA (Pre-Parcial)Carol Julieth Lopez Castillo0% (1)
- DESHIDRATACIÓNDocumento10 pagineDESHIDRATACIÓNLuis TrejoNessuna valutazione finora
- Gases en MinaDocumento87 pagineGases en Minaluisantonio2Nessuna valutazione finora
- Análisis Cuantitativo de Analitos Mediante El Uso de Curvas de CalibraciónDocumento17 pagineAnálisis Cuantitativo de Analitos Mediante El Uso de Curvas de CalibraciónDavid S. ArnedoNessuna valutazione finora
- Agitador MagnéticoDocumento10 pagineAgitador Magnéticoyenny100% (1)
- Monografía - Chavez Leiva Mariano - FacDocumento94 pagineMonografía - Chavez Leiva Mariano - FacCM GetzeNessuna valutazione finora
- Calculo Estructural Del DesarenadorDocumento5 pagineCalculo Estructural Del DesarenadorRenato Enriquez100% (1)
- ANASISSSDocumento2 pagineANASISSSivan100% (2)
- Infografia 3Documento1 paginaInfografia 3carolinaNessuna valutazione finora
- Teoría IrregularidadesDocumento9 pagineTeoría IrregularidadesJairo Alexander ChocontáNessuna valutazione finora
- Tablas R134aDocumento6 pagineTablas R134aCamiloRamirezSanabria0% (1)
- Ochoa Vazquez NoeDocumento91 pagineOchoa Vazquez NoeHobbyNessuna valutazione finora
- El Agua y Su Influencia en La ConstrucciónDocumento8 pagineEl Agua y Su Influencia en La ConstrucciónYeseli Karina Nolasco SifuentesNessuna valutazione finora
- Evolución Cuenca de Falcon GeologiaDocumento34 pagineEvolución Cuenca de Falcon GeologiaJosdelly Daniela Ordaz Ledezma100% (1)
- Rombo de Seguridad y Otros DatosDocumento3 pagineRombo de Seguridad y Otros DatosKaren Chavarro ChauxNessuna valutazione finora
- Boletín 3 15-16Documento2 pagineBoletín 3 15-16Li0% (1)
- Puesta en Marcha y Optimización de Una Planta de Endulzamiento de GasDocumento7 paginePuesta en Marcha y Optimización de Una Planta de Endulzamiento de GasPedro CañizalesNessuna valutazione finora
- Mantenimiento PredictivoDocumento14 pagineMantenimiento PredictivoWENCESLAO VALLEJONessuna valutazione finora
- Dossier TECNOLOGÍA DE PRODUCCIÓN INDUSTRIALDocumento243 pagineDossier TECNOLOGÍA DE PRODUCCIÓN INDUSTRIALrodolf100% (3)
- Cómo Hacer Que El Corazón de Cristal Bórax Día de San ValentínDocumento4 pagineCómo Hacer Que El Corazón de Cristal Bórax Día de San ValentínMarcela Montes ChinchillaNessuna valutazione finora