Potrebbero piacerti anche
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Traditional Healers in South AfricaDocumento8 pagineTraditional Healers in South AfricaTraditional Healer100% (1)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Stroke:: What Should Nurses Need To Know About Stroke?Documento68 pagineStroke:: What Should Nurses Need To Know About Stroke?Suci Aning TNessuna valutazione finora
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Etiology and Diagnosis of Prerenal Disease and Acute Tubular Necrosis in Acute Kidney Injury in Adults - UpToDateDocumento22 pagineEtiology and Diagnosis of Prerenal Disease and Acute Tubular Necrosis in Acute Kidney Injury in Adults - UpToDatecuentaparatrabajosdelau10Nessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- EO 70 S. 2021 - Movement of Pigs and Pork Products ASF-DSDocumento8 pagineEO 70 S. 2021 - Movement of Pigs and Pork Products ASF-DSJamela SerutNessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- WSN 78 (2017) 193-198Documento6 pagineWSN 78 (2017) 193-198Santiago López BuitragoNessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Class 8 - Hand Shao-Yang San-Jiao ChannelDocumento89 pagineClass 8 - Hand Shao-Yang San-Jiao ChannelFrancisco VilaróNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
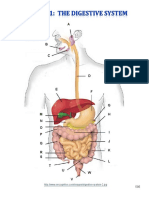
- Digestive System Charts 1Documento14 pagineDigestive System Charts 1api-306762527Nessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Phlegmon: by - Dharmendra Singh Group No. - 317Documento15 paginePhlegmon: by - Dharmendra Singh Group No. - 317Mouna Jomaa DhifaouiNessuna valutazione finora
- Jpog August 2013 IdDocumento51 pagineJpog August 2013 IdHasyim PurwadiNessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Daily Practice Sheet - 105 Molecular Basis of Inheritance: Biomentors Classes Online, Mumbai Date - 27 September 2018Documento6 pagineDaily Practice Sheet - 105 Molecular Basis of Inheritance: Biomentors Classes Online, Mumbai Date - 27 September 2018Sushree DeepaliNessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Anestesi EpiduralDocumento2 pagineAnestesi EpiduralShazean ShahNessuna valutazione finora
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Formulating An Effective Response - A Structured ApproachDocumento22 pagineFormulating An Effective Response - A Structured ApproachMelo MedecieloNessuna valutazione finora
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- TaxonomyDocumento71 pagineTaxonomyNabeel Uddin100% (1)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Deep BretahingDocumento4 pagineDeep Bretahingagustina ika100% (1)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- TRS601 Vocabulary ListDocumento5 pagineTRS601 Vocabulary ListNgo Van Hung (K17 HCM)Nessuna valutazione finora
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- One Health ConceptDocumento2 pagineOne Health ConceptYume AkamaiNessuna valutazione finora
- Post Graduate ClinicsDocumento113 paginePost Graduate ClinicsSnehal PatilNessuna valutazione finora
- Addressing Dry Skin, Acne, Pigmentation & WrinklesDocumento26 pagineAddressing Dry Skin, Acne, Pigmentation & WrinklesMucharla Praveen KumarNessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- CL Septembre Grandma RemediesDocumento8 pagineCL Septembre Grandma RemediesMaria BenessalahNessuna valutazione finora
- Posted by Ned Haluzan, 0 Comments Labels:: Air PollutionDocumento7 paginePosted by Ned Haluzan, 0 Comments Labels:: Air PollutionPei Jue TanNessuna valutazione finora
- Announcement Sinas IDAI - Draft 07Documento5 pagineAnnouncement Sinas IDAI - Draft 07Hendra JayaNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- Script For OrientationDocumento2 pagineScript For OrientationAngela CudiaNessuna valutazione finora
- Bài Kiểm Tra Thường Xuyên Lần 2Documento3 pagineBài Kiểm Tra Thường Xuyên Lần 2Hoctienganh HhNessuna valutazione finora
- Shafers Textbook of Oral Pathology 9Th Edition William G Shafer All ChapterDocumento67 pagineShafers Textbook of Oral Pathology 9Th Edition William G Shafer All Chaptermary.graham20483% (6)
- MVG09Documento4 pagineMVG09Sondos HarbiehNessuna valutazione finora
- Lec 4-Toxicity and Identification TestDocumento46 pagineLec 4-Toxicity and Identification TestShoaib MuhammadNessuna valutazione finora
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- OB TriageDocumento36 pagineOB TriagerelinNessuna valutazione finora
- Gynecological NursingDocumento329 pagineGynecological Nursingsharon ocharaNessuna valutazione finora
- SunCoBe ShampooDocumento24 pagineSunCoBe ShampooDong YulNessuna valutazione finora
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)