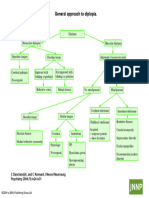
Potrebbero piacerti anche
- Antioxidants 11 00692Documento19 pagineAntioxidants 11 00692Gadisa Aulia PratamiNessuna valutazione finora
- Metabolism FeDocumento17 pagineMetabolism Feeduardo sosaNessuna valutazione finora
- Oxidative Stress and The Homeodynamics of Iron MetabolismDocumento40 pagineOxidative Stress and The Homeodynamics of Iron MetabolismRuwi KavindyaNessuna valutazione finora
- Medicinal Chemistry PortfolioDocumento17 pagineMedicinal Chemistry PortfolioTan Meng ChuNessuna valutazione finora
- Mechanisms of Iron Loading and Toxicity: Gregory J. AndersonDocumento4 pagineMechanisms of Iron Loading and Toxicity: Gregory J. Andersonikhsan07Nessuna valutazione finora
- NIH Public Access: Transition Metals and Mitochondrial Metabolism in The HeartDocumento17 pagineNIH Public Access: Transition Metals and Mitochondrial Metabolism in The HeartAlix AliNessuna valutazione finora
- Mechanisms Controlling Cellular - Nature Ncoa4 Ekim23Documento23 pagineMechanisms Controlling Cellular - Nature Ncoa4 Ekim23kocagerahmetNessuna valutazione finora
- Metabolisme IronDocumento15 pagineMetabolisme IronERNI FORWATYNessuna valutazione finora
- Asian Paci Fic Journal of Tropical Medicine: Iron Dysregulation in Beta-ThalassemiaDocumento9 pagineAsian Paci Fic Journal of Tropical Medicine: Iron Dysregulation in Beta-ThalassemiafitriahendricoNessuna valutazione finora
- Lecture 15 PDFDocumento20 pagineLecture 15 PDFSrramNessuna valutazione finora
- Copper and Iron Homeostasis in Plants: The Challenges of Oxidative StressDocumento14 pagineCopper and Iron Homeostasis in Plants: The Challenges of Oxidative StressEddard StarkNessuna valutazione finora
- Investigación Ácido ÚricoDocumento27 pagineInvestigación Ácido ÚricoAnonymous K8fT6HNessuna valutazione finora
- Heavy Metal Induced Oxidative Stress & Its Possible Reversal by Chelation TherapyDocumento23 pagineHeavy Metal Induced Oxidative Stress & Its Possible Reversal by Chelation TherapyArnab BiswasNessuna valutazione finora
- Che - 91911 Significance of Cu2+Documento1 paginaChe - 91911 Significance of Cu2+Jo StandleyNessuna valutazione finora
- Processes: Metal Ions, Metal Chelators and Metal Chelating Assay As Antioxidant MethodDocumento16 pagineProcesses: Metal Ions, Metal Chelators and Metal Chelating Assay As Antioxidant Methodwow tryNessuna valutazione finora
- Human Iron Metabolism - WikipediaDocumento37 pagineHuman Iron Metabolism - WikipediaMaysaa AlashkarNessuna valutazione finora
- Experimental Eye Research: M. Goralska, J. Ferrell, J. Harned, M. Lall, S. Nagar, L.N. Fleisher, M.C. McgahanDocumento12 pagineExperimental Eye Research: M. Goralska, J. Ferrell, J. Harned, M. Lall, S. Nagar, L.N. Fleisher, M.C. McgahanPriscila Verduzco MartínezNessuna valutazione finora
- Metal Homeostasis: Advanced Article F. Wayne Outten and Benjamin S. TwiningDocumento10 pagineMetal Homeostasis: Advanced Article F. Wayne Outten and Benjamin S. TwiningazzaassNessuna valutazione finora
- Sidd Iq Que 2012Documento18 pagineSidd Iq Que 2012Joanne RoxanneNessuna valutazione finora
- Superoxides Dismutase, A Ancient Enzym and New InsightDocumento11 pagineSuperoxides Dismutase, A Ancient Enzym and New InsightRizkiantoNessuna valutazione finora
- Unit - I Bio InorganicDocumento16 pagineUnit - I Bio Inorganicharirajans71Nessuna valutazione finora
- Topic - 6 - QR of D-ElementsDocumento14 pagineTopic - 6 - QR of D-Elementststtwa LyoaNessuna valutazione finora
- Iron MetabolismDocumento21 pagineIron MetabolismInês FerreiraNessuna valutazione finora
- Iron in FoodDocumento22 pagineIron in FoodMarshal GrahamNessuna valutazione finora
- Hepcidin Knockout Mice Fed With Iron Rich DietDocumento9 pagineHepcidin Knockout Mice Fed With Iron Rich DietEduardo Henrique de Sousa SalvinoNessuna valutazione finora
- 1-Principles in Bioinorganic Chemistry Traducir.Documento6 pagine1-Principles in Bioinorganic Chemistry Traducir.CharlieWallNessuna valutazione finora
- Metal Ions in Biological Systems PDFDocumento4 pagineMetal Ions in Biological Systems PDFVigyan PravahaNessuna valutazione finora
- Biochimica Et Biophysica Acta: Tomas Ganz, Elizabeta NemethDocumento10 pagineBiochimica Et Biophysica Acta: Tomas Ganz, Elizabeta NemethAnonymous oUlErgOINessuna valutazione finora
- MetalloenzymesDocumento5 pagineMetalloenzymesAzabo KennedyNessuna valutazione finora
- TRACE MineralsDocumento18 pagineTRACE MineralsKipchirchir AbednegoNessuna valutazione finora
- Are Fe-Based Stenting Materials Biocompatible A Critical Review of in Vitro and in Vivo StudiesDocumento16 pagineAre Fe-Based Stenting Materials Biocompatible A Critical Review of in Vitro and in Vivo StudiesManas Ranjan SahuNessuna valutazione finora
- Metabolites 12 00609 v2Documento11 pagineMetabolites 12 00609 v2Ionela MariaNessuna valutazione finora
- Activity-Based Sensing Fluorescent Probes For Iron in Biological SystemsDocumento6 pagineActivity-Based Sensing Fluorescent Probes For Iron in Biological SystemsVijay LouNessuna valutazione finora
- The Role of Copper As A Modifier of Lipid Metabolism: January 2013Documento23 pagineThe Role of Copper As A Modifier of Lipid Metabolism: January 2013Leni LismayantiNessuna valutazione finora
- Iron BalanceDocumento7 pagineIron BalancegigibesiNessuna valutazione finora
- Copper DetDocumento12 pagineCopper DetInna popaNessuna valutazione finora
- Jurowski2014 Article BiologicalConsequencesOfZincDeDocumento11 pagineJurowski2014 Article BiologicalConsequencesOfZincDevtimbangjrNessuna valutazione finora
- Balance de HierroDocumento13 pagineBalance de HierroWalter Huacani HuamaniNessuna valutazione finora
- The Cellular Effect of Lead Poisoning and Its Clinical PictureDocumento8 pagineThe Cellular Effect of Lead Poisoning and Its Clinical PictureRina OktavianaNessuna valutazione finora
- Alleviation of Iron Deficiency in Plants, Animals and Humans Through Biofortification A Detailed ReviewDocumento8 pagineAlleviation of Iron Deficiency in Plants, Animals and Humans Through Biofortification A Detailed ReviewInternational Journal of Innovative Science and Research TechnologyNessuna valutazione finora
- Ketidakseimbangan Cu Dan ZNDocumento18 pagineKetidakseimbangan Cu Dan ZNMuthia FadhilahNessuna valutazione finora
- 03 Iron Transport Proteins Gateways of Cellular and SystemicDocumento9 pagine03 Iron Transport Proteins Gateways of Cellular and SystemicMatheus SouzaNessuna valutazione finora
- Iron and Copper Metabolism: Miguel Arredondo, Marco T. Nu N EzDocumento15 pagineIron and Copper Metabolism: Miguel Arredondo, Marco T. Nu N EzkonyekNessuna valutazione finora
- Mechanisms of Cell InjuryDocumento68 pagineMechanisms of Cell InjuryAritra HalderNessuna valutazione finora
- Electrons Redox Scavenging Free Radicals (1) (2) : CopperDocumento9 pagineElectrons Redox Scavenging Free Radicals (1) (2) : CopperTowhid HasanNessuna valutazione finora
- Metalloregulatory Proteins: Advanced Article Deborah B. ZambleDocumento10 pagineMetalloregulatory Proteins: Advanced Article Deborah B. ZambleazzaassNessuna valutazione finora
- Iron Physiology of IronDocumento7 pagineIron Physiology of IronAnna PodsiadłoNessuna valutazione finora
- FerritinDocumento15 pagineFerritinericmeitei100% (1)
- Bioinorg Anic Chemist Ry: Bulk Elements ( Organic') - O, H, C, N, S, PDocumento4 pagineBioinorg Anic Chemist Ry: Bulk Elements ( Organic') - O, H, C, N, S, PEllie CarterNessuna valutazione finora
- DissertationDocumento29 pagineDissertationShobha GoswamiNessuna valutazione finora
- Iron Biology, Immunology, Aging, and Obesity: Four Fields Connected by The Small Peptide Hormone HepcidinDocumento16 pagineIron Biology, Immunology, Aging, and Obesity: Four Fields Connected by The Small Peptide Hormone HepcidinSagita DarmasariNessuna valutazione finora
- Walters, Esfandi, Tsopmo - 2018 - Potential of Food Hydrolyzed Proteins and Peptides To Chelate Iron or Calcium and Enhance Their Absorp-AnnotatedDocumento17 pagineWalters, Esfandi, Tsopmo - 2018 - Potential of Food Hydrolyzed Proteins and Peptides To Chelate Iron or Calcium and Enhance Their Absorp-AnnotatedDiễm SươngNessuna valutazione finora
- Tinjauan Pustaka: Hubungan Besi Dan Produksi SitokinDocumento10 pagineTinjauan Pustaka: Hubungan Besi Dan Produksi SitokinAulia PuspitaNessuna valutazione finora
- Níveis de FerroDocumento13 pagineNíveis de FerroCarla FernandaNessuna valutazione finora
- Hepcidin, A Key Regulator of Iron Metabolism and Mediator of Anemia of InflammationDocumento6 pagineHepcidin, A Key Regulator of Iron Metabolism and Mediator of Anemia of InflammationCesar Josué GarcíaNessuna valutazione finora
- Chemistry of Minerals-IIDocumento29 pagineChemistry of Minerals-IIȘudîpțo ȘhăhîdNessuna valutazione finora
- Chemistry of Minerals-IIDocumento29 pagineChemistry of Minerals-IIȘudîpțo ȘhăhîdNessuna valutazione finora
- Metal-Deficient SOD1 in Amyotrophic Lateral Sclerosis: ReviewDocumento7 pagineMetal-Deficient SOD1 in Amyotrophic Lateral Sclerosis: ReviewLuz Eneida CabreraNessuna valutazione finora
- Essential Non Essential Metals and Human HealthDocumento50 pagineEssential Non Essential Metals and Human HealthLi LissNessuna valutazione finora
- Diplopia - Eye Disorders - MSD Manual Professional Edition-TableDocumento3 pagineDiplopia - Eye Disorders - MSD Manual Professional Edition-TableApostolos T.Nessuna valutazione finora
- Depression SerotoninDocumento5 pagineDepression SerotoninidjacobsNessuna valutazione finora
- Diplopia - Eye Disorders - MSD Manual Professional EditionDocumento5 pagineDiplopia - Eye Disorders - MSD Manual Professional EditionApostolos T.Nessuna valutazione finora
- Jmf.2020.0137-Genoderma in PDDocumento8 pagineJmf.2020.0137-Genoderma in PDApostolos T.Nessuna valutazione finora
- Diplopia-jnnp-2004-December-75-suppl 4-Iv24Documento1 paginaDiplopia-jnnp-2004-December-75-suppl 4-Iv24Apostolos T.Nessuna valutazione finora
- PAIN IN PARKINSON'S DISEASE-50 Cases (2008)Documento4 paginePAIN IN PARKINSON'S DISEASE-50 Cases (2008)Apostolos T.Nessuna valutazione finora
- Trigemina Neuralgia Guidelines - 2021 - V4Documento21 pagineTrigemina Neuralgia Guidelines - 2021 - V4Apostolos T.Nessuna valutazione finora
- MiorelDocumento1 paginaMiorelApostolos T.Nessuna valutazione finora
- PAINWeek Journal Vol 7, Q1Documento78 paginePAINWeek Journal Vol 7, Q1Apostolos T.Nessuna valutazione finora
- Assessment of Efficacy and Safety of Eslicarbazepine Acetate For The Treatment of Trigeminal NeuralgiaDocumento9 pagineAssessment of Efficacy and Safety of Eslicarbazepine Acetate For The Treatment of Trigeminal NeuralgiaApostolos T.Nessuna valutazione finora
- Pain: Current Understanding of Assessment, Management, and TreatmentsDocumento101 paginePain: Current Understanding of Assessment, Management, and TreatmentsNational Pharmaceutical Council100% (8)
- Adverse Effects of Antiepileptic Drugs (2010)Documento8 pagineAdverse Effects of Antiepileptic Drugs (2010)Apostolos T.Nessuna valutazione finora
- PAINWeek Journal Vol 5, Q1Documento62 paginePAINWeek Journal Vol 5, Q1Apostolos T.Nessuna valutazione finora
- Vimpat EngDocumento148 pagineVimpat EngApostolos T.Nessuna valutazione finora
- NIH Public Access: Author ManuscriptDocumento48 pagineNIH Public Access: Author ManuscriptApostolos T.Nessuna valutazione finora
- Thursday, October 29, 2020: Cony 2020 Virtual Congress Scientific Program (Subject To Changes)Documento24 pagineThursday, October 29, 2020: Cony 2020 Virtual Congress Scientific Program (Subject To Changes)Apostolos T.Nessuna valutazione finora
- Fibromyalgia & Trazodone+Pregabalin 2011Documento9 pagineFibromyalgia & Trazodone+Pregabalin 2011Apostolos T.Nessuna valutazione finora
- ITCHDocumento18 pagineITCHApostolos T.Nessuna valutazione finora
- The Status of Diagnosis and Treatment To Intracranial Hypotension (2017)Documento8 pagineThe Status of Diagnosis and Treatment To Intracranial Hypotension (2017)Apostolos T.Nessuna valutazione finora
- Cephalalgia ReviewDocumento25 pagineCephalalgia ReviewApostolos T.Nessuna valutazione finora
- Diagnosing Secondary Headaches (Practical Neurology) (2020 - 05)Documento5 pagineDiagnosing Secondary Headaches (Practical Neurology) (2020 - 05)Apostolos T.Nessuna valutazione finora
- Functional Magnetic Resonance Imaging in Episodic Cluster Headache (2009)Documento4 pagineFunctional Magnetic Resonance Imaging in Episodic Cluster Headache (2009)Apostolos T.Nessuna valutazione finora
- Neuroimaging in Cluster Headache and Other Trigeminal Autonomic Cephalalgias (2012)Documento10 pagineNeuroimaging in Cluster Headache and Other Trigeminal Autonomic Cephalalgias (2012)Apostolos T.Nessuna valutazione finora
- Understanding Cluster Headache Using MRI (2020)Documento12 pagineUnderstanding Cluster Headache Using MRI (2020)Apostolos T.Nessuna valutazione finora
- A Role For FMRI in Optimizing CNS Drug DDocumento14 pagineA Role For FMRI in Optimizing CNS Drug DApostolos T.Nessuna valutazione finora
- Setup: Original Language (English) Your Language TranslationDocumento25 pagineSetup: Original Language (English) Your Language TranslationApostolos T.Nessuna valutazione finora
- Overview of Standardized HRV Assessment Protocol - 2Documento7 pagineOverview of Standardized HRV Assessment Protocol - 2Apostolos T.Nessuna valutazione finora
- Adult EpilepsyDocumento14 pagineAdult Epilepsyammarhafez78Nessuna valutazione finora
- Metals: Iron, Aging, and NeurodegenerationDocumento23 pagineMetals: Iron, Aging, and NeurodegenerationApostolos T.Nessuna valutazione finora
- Cerebral Amyloid Angiopathy A Systematic Review 2011Documento9 pagineCerebral Amyloid Angiopathy A Systematic Review 2011Jaime Arevalo PanduroNessuna valutazione finora
- Stem Cell Treatment Alzheimer's Disease - ASCI - Asian Stem Cell InstituteDocumento39 pagineStem Cell Treatment Alzheimer's Disease - ASCI - Asian Stem Cell Institutea73072Nessuna valutazione finora
- Encyclopedia of Biological Chemistry - Vol - 4Documento503 pagineEncyclopedia of Biological Chemistry - Vol - 4joshigauta100% (1)
- The Proteomic Response To Metabolic Stress and Cellular Dysfunction in Relation To Alzheimer's DiseaseDocumento312 pagineThe Proteomic Response To Metabolic Stress and Cellular Dysfunction in Relation To Alzheimer's DiseaseSpruce ApparelsNessuna valutazione finora
- Alzheimer's Disease: From The Bench To The ClinicDocumento37 pagineAlzheimer's Disease: From The Bench To The Clinichoyvin100% (1)
- Biology Presentation Alzheimer'sDocumento14 pagineBiology Presentation Alzheimer'skkathrynanna100% (4)
- CerebrolysinDocumento32 pagineCerebrolysinRena KallergiNessuna valutazione finora
- Nishi's QB - PhathoDocumento42 pagineNishi's QB - PhathofrabziNessuna valutazione finora
- Plasma Ab42 and Ab40 As Markers of CognitiveDocumento5 paginePlasma Ab42 and Ab40 As Markers of CognitivePetru BalaceanuNessuna valutazione finora
- ბეტა ამილოიდი და ტაუ ცილა ალცჰაიმერის დაავადების პათოგენეზშიDocumento1 paginaბეტა ამილოიდი და ტაუ ცილა ალცჰაიმერის დაავადების პათოგენეზშიEMD GROUPNessuna valutazione finora
- Alzheimer's - Alzheimer's Disease - Kaj Blennow, Mony J de Leon, Henrik ZeterbergDocumento17 pagineAlzheimer's - Alzheimer's Disease - Kaj Blennow, Mony J de Leon, Henrik ZeterbergCatinean DanielaNessuna valutazione finora
- Pathophysiology of Alzheimer's Disease: Neuroimaging Clinics of North America December 2005Documento28 paginePathophysiology of Alzheimer's Disease: Neuroimaging Clinics of North America December 2005AnisNessuna valutazione finora
- Post-Translational Modifications of Proteins: Acetylcholinesterase As A Model SystemDocumento17 paginePost-Translational Modifications of Proteins: Acetylcholinesterase As A Model SystemSergio AraujoNessuna valutazione finora
- Alzheimer's DiseaseDocumento29 pagineAlzheimer's DiseaseKeVz Nicdao100% (1)
- THE GENETICS OF ALZHEIMERS DISEASE. PPTDocumento24 pagineTHE GENETICS OF ALZHEIMERS DISEASE. PPTBenjaminVasileniucNessuna valutazione finora
- Atlas of Alzheimer's DiseaseDocumento159 pagineAtlas of Alzheimer's DiseaseDanStoica83% (6)
- Review HMG-CoA ReductaseDocumento10 pagineReview HMG-CoA ReductaseFrancisco TapiaNessuna valutazione finora
- Drugs in Clinical Trials For Alzheimer'sDocumento40 pagineDrugs in Clinical Trials For Alzheimer'sEdward Vega LaraNessuna valutazione finora
- Yoshikuni Mizuno, Abraham Fisher, Israel Hanin - Mapping The Progress of Alzheimer's and Parkinson's Disease (Advances in Behavioral Biology) (2002) PDFDocumento588 pagineYoshikuni Mizuno, Abraham Fisher, Israel Hanin - Mapping The Progress of Alzheimer's and Parkinson's Disease (Advances in Behavioral Biology) (2002) PDFRobert Cristian GodeanuNessuna valutazione finora
- Alzheimer: Yovita Setiady (406138120) Nieta Hardiyanti (406148067) Meilisa Tiffani (406148069)Documento29 pagineAlzheimer: Yovita Setiady (406138120) Nieta Hardiyanti (406148067) Meilisa Tiffani (406148069)Nita nuriasNessuna valutazione finora
- Plasticity As A Therapeutic Target For Improving Cognition and Behavior in Down Syndrome.Documento34 paginePlasticity As A Therapeutic Target For Improving Cognition and Behavior in Down Syndrome.EdsonDouradoMenezesNessuna valutazione finora
- Pinweha2008 PDFDocumento5 paginePinweha2008 PDFDr. Kaushal Kishor SharmaNessuna valutazione finora
- A Critical Appraisal of Amyloid-β-targeting Therapies for Alzheimer Disease 2019 ReviewDocumento16 pagineA Critical Appraisal of Amyloid-β-targeting Therapies for Alzheimer Disease 2019 Review畏Nessuna valutazione finora
- The Pathophysiology of Alzheimer'S Disease and Directions in TreatmentDocumento15 pagineThe Pathophysiology of Alzheimer'S Disease and Directions in TreatmentEnerolisa ParedesNessuna valutazione finora
- Transgenic ValidationDocumento22 pagineTransgenic ValidationSyama J.S33% (6)
- Beta-Secretase: Structure, Function, and EvolutionDocumento18 pagineBeta-Secretase: Structure, Function, and EvolutionAlix AliNessuna valutazione finora
- Althimer's ResearchDocumento99 pagineAlthimer's Researchdeemoney3Nessuna valutazione finora
- Management of Oxidative Stress and Other PathologiDocumento23 pagineManagement of Oxidative Stress and Other PathologiyusufNessuna valutazione finora
- Understanding Alzheimers DiseaseDocumento494 pagineUnderstanding Alzheimers Diseasegezedka100% (3)
- Cia 2 347Documento15 pagineCia 2 347AnisNessuna valutazione finora
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDa EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouValutazione: 4.5 su 5 stelle4.5/5 (62)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDa EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityValutazione: 4 su 5 stelle4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDa EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisValutazione: 3.5 su 5 stelle3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDa EverandTales from Both Sides of the Brain: A Life in NeuroscienceValutazione: 3 su 5 stelle3/5 (18)
- Gut: the new and revised Sunday Times bestsellerDa EverandGut: the new and revised Sunday Times bestsellerValutazione: 4 su 5 stelle4/5 (392)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDa EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedValutazione: 4 su 5 stelle4/5 (11)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDa EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceValutazione: 4.5 su 5 stelle4.5/5 (516)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Da EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Valutazione: 4 su 5 stelle4/5 (378)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDa Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessValutazione: 4 su 5 stelle4/5 (33)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDa EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionValutazione: 4 su 5 stelle4/5 (811)
- Inside of a Dog: What Dogs See, Smell, and KnowDa EverandInside of a Dog: What Dogs See, Smell, and KnowValutazione: 4 su 5 stelle4/5 (390)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDa EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsValutazione: 4 su 5 stelle4/5 (5)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDa EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeNessuna valutazione finora
- Human: The Science Behind What Makes Your Brain UniqueDa EverandHuman: The Science Behind What Makes Your Brain UniqueValutazione: 3.5 su 5 stelle3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDa EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperValutazione: 4.5 su 5 stelle4.5/5 (15)
- Good Without God: What a Billion Nonreligious People Do BelieveDa EverandGood Without God: What a Billion Nonreligious People Do BelieveValutazione: 4 su 5 stelle4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDa EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNessuna valutazione finora
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDa EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondValutazione: 4 su 5 stelle4/5 (4)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomDa EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomValutazione: 4 su 5 stelle4/5 (215)
- Who's in Charge?: Free Will and the Science of the BrainDa EverandWho's in Charge?: Free Will and the Science of the BrainValutazione: 4 su 5 stelle4/5 (65)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDa EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesValutazione: 4.5 su 5 stelle4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDa EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldValutazione: 4.5 su 5 stelle4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineDa EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineValutazione: 4 su 5 stelle4/5 (17)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDa EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemValutazione: 4.5 su 5 stelle4.5/5 (115)
- The Invention of Tomorrow: A Natural History of ForesightDa EverandThe Invention of Tomorrow: A Natural History of ForesightValutazione: 4.5 su 5 stelle4.5/5 (5)