Potrebbero piacerti anche
- Sistema Del ComplementoDocumento19 pagineSistema Del ComplementoSergio David Salazar Lemus100% (1)
- PreLab 6,7Documento7 paginePreLab 6,7Ana Ceto BritoNessuna valutazione finora
- Bombas de protones en levaduras y los efectos de desacoplantes e inhibidoresDocumento10 pagineBombas de protones en levaduras y los efectos de desacoplantes e inhibidoresAnnie Villegas0% (1)
- Mar-11-Extracción de Adn de Células VegetalesDocumento10 pagineMar-11-Extracción de Adn de Células VegetalesStephany Mariela Espinoza SachaNessuna valutazione finora
- GPC VIH Acuerdo Ministerial05!07!2019Documento156 pagineGPC VIH Acuerdo Ministerial05!07!2019Kate CastañoNessuna valutazione finora
- Tarjeta navidad Los SimuladoresDocumento1 paginaTarjeta navidad Los SimuladoresPedroCordovageNessuna valutazione finora
- Trichuris TrichiuraDocumento4 pagineTrichuris TrichiuraJuan MartinezNessuna valutazione finora
- Ppractica 1. Principales Tecnicas en El Manejo de AnimalesDocumento9 paginePpractica 1. Principales Tecnicas en El Manejo de AnimalesBRANDEN EDUARDO PAISANO ALVAREZNessuna valutazione finora
- Bioseguridad en El Laboratorio de MicologíaDocumento13 pagineBioseguridad en El Laboratorio de MicologíaDario Gomez0% (1)
- Oxidación y Sindrome de Dress Expo 1 FarmacoDocumento14 pagineOxidación y Sindrome de Dress Expo 1 FarmacoMEDICINA PARA TODOSNessuna valutazione finora
- Estructura Del Citocromo P450Documento3 pagineEstructura Del Citocromo P450Juniors SalvatierraNessuna valutazione finora
- Chilomastix MesniliDocumento3 pagineChilomastix MesniliAngieNessuna valutazione finora
- Dilución en SerieDocumento2 pagineDilución en SerieAlejandro A. Burgos50% (2)
- Observación de Protozoarios Estudiados en ClaseDocumento14 pagineObservación de Protozoarios Estudiados en ClaseCésar ArteagaNessuna valutazione finora
- Protistas Del Clado Parabasalia en TermitaDocumento7 pagineProtistas Del Clado Parabasalia en TermitaAlan SalasNessuna valutazione finora
- Citoesqueleto ForoDocumento39 pagineCitoesqueleto ForoClaudia Montes GuerraNessuna valutazione finora
- Termociclador Convencional (PCR)Documento8 pagineTermociclador Convencional (PCR)Nora Quispe CamaticonaNessuna valutazione finora
- Solanum LycopersicumDocumento13 pagineSolanum LycopersicumBrian Anthony Suarez FernandezNessuna valutazione finora
- Practica Reconocimiento de Enzimas 6Documento9 paginePractica Reconocimiento de Enzimas 6Leonardo Francisco Sanchez GuerreroNessuna valutazione finora
- Marco Teorico Sobre LeishmaniasisDocumento8 pagineMarco Teorico Sobre LeishmaniasisSG SimónNessuna valutazione finora
- Previo 3 - CoprocultivoDocumento5 paginePrevio 3 - CoprocultivoMelissa GalanNessuna valutazione finora
- Práctica de Laboratorio Cilios y FlagelosDocumento7 paginePráctica de Laboratorio Cilios y FlagelosMarilyn Guamuch García0% (1)
- Guia de Halogenuros de AlquiloDocumento20 pagineGuia de Halogenuros de AlquiloLuis Carlos Andrade100% (1)
- Aspectos Sociales de La ParasitologiaDocumento17 pagineAspectos Sociales de La ParasitologiaHaydee MartinezNessuna valutazione finora
- Dientamoeba FragilisDocumento4 pagineDientamoeba Fragilisclaudiap2802Nessuna valutazione finora
- Grupos funcionales orgánicos vitalesDocumento22 pagineGrupos funcionales orgánicos vitalesEmmanuel Pacheco100% (1)
- Identificación de Azúcares y Almidones - LugolDocumento4 pagineIdentificación de Azúcares y Almidones - LugolLupita AlanNessuna valutazione finora
- Técnicas histológicasDocumento11 pagineTécnicas histológicasAníbal CINessuna valutazione finora
- TP Nº10. CarbohidratosDocumento5 pagineTP Nº10. CarbohidratosJana Vecchio RomeroNessuna valutazione finora
- Micro FinalDocumento225 pagineMicro FinalJenny AndreinaNessuna valutazione finora
- Reproduccion BacterianaDocumento6 pagineReproduccion Bacterianajngvadvp0% (1)
- Importancia de Las Aminas en AnimalesDocumento16 pagineImportancia de Las Aminas en AnimalesCitlalli Arcadia Orozco100% (1)
- Clastridium Botulinum - Botulismo Alimenticio en HumanosDocumento15 pagineClastridium Botulinum - Botulismo Alimenticio en HumanosCamila Rios RamosNessuna valutazione finora
- Furanos y FenolesDocumento10 pagineFuranos y FenolesAmado Quispe QuispeNessuna valutazione finora
- Puntos de RestricciónDocumento20 paginePuntos de RestricciónLuis Copalcua LopantziNessuna valutazione finora
- Chaperoninas ListoDocumento23 pagineChaperoninas ListoDuvan Zavaleta Bazan100% (1)
- Practica 1 - Observación Simple de Hongos y LevadurasDocumento3 paginePractica 1 - Observación Simple de Hongos y LevadurasLaura Recalde25% (4)
- Protozoos Flagelados (Zoomastigophoros Intestinales)Documento25 pagineProtozoos Flagelados (Zoomastigophoros Intestinales)José Gustavo Gaitán FloresNessuna valutazione finora
- Permeabilidad LaboratorioDocumento11 paginePermeabilidad LaboratoriomelyNessuna valutazione finora
- Microscopía y técnicas histológicasDocumento6 pagineMicroscopía y técnicas histológicasCabito CatalanNessuna valutazione finora
- UreaplasmaDocumento2 pagineUreaplasmaMayi Jimenez0% (1)
- Guia de Alcoholes y FenolesDocumento3 pagineGuia de Alcoholes y FenolesPaulo Alejandro Araos Brito100% (1)
- Género CorynebacteriumDocumento20 pagineGénero CorynebacteriumYael Yomira Ramos TatajeNessuna valutazione finora
- CandidiasisDocumento4 pagineCandidiasisaraalmaguerNessuna valutazione finora
- Guanarito B9Documento5 pagineGuanarito B9Karethn YepezNessuna valutazione finora
- Cuadro Características EnterobacteriasDocumento3 pagineCuadro Características EnterobacteriasCesar Nicolas SosaNessuna valutazione finora
- Caracterización Del Genero PseudomonasDocumento3 pagineCaracterización Del Genero PseudomonasAbiram PardoNessuna valutazione finora
- Cuestionario LiquenesDocumento6 pagineCuestionario LiquenesNahomy AhedoNessuna valutazione finora
- Reconocimiento Células SanguíneasDocumento5 pagineReconocimiento Células SanguíneasLucia Arriaga DNessuna valutazione finora
- Informacion de IsomeriaDocumento9 pagineInformacion de Isomeriaximena rodriguez cadenaNessuna valutazione finora
- Biotecnología RojaDocumento3 pagineBiotecnología RojaCarlos SerranoNessuna valutazione finora
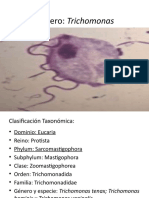
- Género TrichomonasDocumento57 pagineGénero Trichomonasandroth alonso ruizNessuna valutazione finora
- CRUCIGRAMA 2 AlumnosDocumento1 paginaCRUCIGRAMA 2 Alumnosdan50% (2)
- Amiloidosis PDFDocumento7 pagineAmiloidosis PDFluisillo007Nessuna valutazione finora
- Ciclo Celular ProcarióticoDocumento11 pagineCiclo Celular Procarióticoatomo5250% (6)
- Morfologia MicrobianaDocumento9 pagineMorfologia Microbiana.:("*"BLacK BuLLeT"*"):.33% (3)
- RESUMEN#5 Inmunidad Especializada en Las Barreras y en Los Tejidos Con Privilegio InmunitarioDocumento6 pagineRESUMEN#5 Inmunidad Especializada en Las Barreras y en Los Tejidos Con Privilegio InmunitarioJessicaNessuna valutazione finora
- Fosforilación OxidativaDocumento32 pagineFosforilación OxidativaJasonRodriguezGuerraNessuna valutazione finora
- Tripsina InhibidoresDocumento11 pagineTripsina InhibidoresJonathan Andres Ortiz ForeroNessuna valutazione finora
- Normas de bioseguridad en el laboratorioDocumento8 pagineNormas de bioseguridad en el laboratoriodejomiNessuna valutazione finora
- Glucólisis: Oxidación GlucosaDocumento32 pagineGlucólisis: Oxidación GlucosaCe Ga75% (4)
- Toxoplasmosis Evasion - En.esDocumento8 pagineToxoplasmosis Evasion - En.esKatherine Shirley Patiño CopaNessuna valutazione finora
- Mecanismo Inmunologico Del DengueDocumento10 pagineMecanismo Inmunologico Del DengueNadia Alexandra Agurto100% (1)
- Tipos Graficos Act1 U6Documento3 pagineTipos Graficos Act1 U6PedroCordovageNessuna valutazione finora
- Vida UtilDocumento5 pagineVida UtilPedroCordovageNessuna valutazione finora
- Bebida Sanky: análisis de marketingDocumento22 pagineBebida Sanky: análisis de marketingPedroCordovageNessuna valutazione finora
- V 14 N 1 TR 1Documento12 pagineV 14 N 1 TR 1Lennin Alcántara ValleNessuna valutazione finora
- Vancomicina, Pac HemodialisisDocumento5 pagineVancomicina, Pac HemodialisisPedroCordovageNessuna valutazione finora
- Vocabulario InglesDocumento58 pagineVocabulario InglesMiguel Angel Criado AlonsoNessuna valutazione finora
- CRIOAGLUTININASDocumento1 paginaCRIOAGLUTININASPedroCordovageNessuna valutazione finora
- Obtención de Lisina Libre A Partir de Quinua OrgánicaDocumento3 pagineObtención de Lisina Libre A Partir de Quinua OrgánicaPedroCordovage100% (1)
- Etiología y Fisiopatogenia EsquizofreniaDocumento3 pagineEtiología y Fisiopatogenia EsquizofreniaPedroCordovageNessuna valutazione finora
- Agar AgarDocumento4 pagineAgar Agarailyn heidy yanc baylonNessuna valutazione finora
- TBC TripticoDocumento2 pagineTBC TripticoPedroCordovageNessuna valutazione finora
- Tecnovigilancia y FarmacovigilanciaDocumento7 pagineTecnovigilancia y FarmacovigilanciaPedroCordovageNessuna valutazione finora
- Alimentos Composicion y Propiedades AstiasaranDocumento4 pagineAlimentos Composicion y Propiedades AstiasaranPedroCordovageNessuna valutazione finora
- Control Microbiológico Jarina de ChiaDocumento1 paginaControl Microbiológico Jarina de ChiaPedroCordovageNessuna valutazione finora
- Queso ParmesanoDocumento3 pagineQueso ParmesanoPedroCordovage50% (2)
- GlucemiaDocumento14 pagineGlucemiaPedroCordovageNessuna valutazione finora
- Metodo de Kahane5, DenigesDocumento4 pagineMetodo de Kahane5, DenigesAnibal Quispe VqzNessuna valutazione finora
- Gen XpertDocumento8 pagineGen XpertPedroCordovageNessuna valutazione finora
- Agar AgarDocumento4 pagineAgar Agarailyn heidy yanc baylonNessuna valutazione finora
- Antecedentes Del Paso UnoDocumento6 pagineAntecedentes Del Paso UnoPedroCordovageNessuna valutazione finora
- Vivir Mejor FDJ PDFDocumento76 pagineVivir Mejor FDJ PDFAndrea Paz FigueroaNessuna valutazione finora
- CcosmeticoDocumento3 pagineCcosmeticosociedad de servicios farmaceuticos a la comunidadNessuna valutazione finora
- A Bilirrubinas T, D e I - 2018Documento33 pagineA Bilirrubinas T, D e I - 2018PedroCordovageNessuna valutazione finora
- Modelo MalcomDocumento3 pagineModelo MalcomPedroCordovageNessuna valutazione finora
- Alteraciones de Los AlimentosDocumento14 pagineAlteraciones de Los AlimentosLuz Martinez MartinezNessuna valutazione finora
- Requisitos para Inscripción en El CQFDLDocumento2 pagineRequisitos para Inscripción en El CQFDLPedroCordovageNessuna valutazione finora
- cOACHING Y MENTORDocumento3 paginecOACHING Y MENTORPedroCordovageNessuna valutazione finora
- Costes calidad empresaDocumento17 pagineCostes calidad empresaHenry TaboadaNessuna valutazione finora
- Infeccion Por El Virus de Inmunodeficiencia HumanaDocumento36 pagineInfeccion Por El Virus de Inmunodeficiencia Humanaalondra moralesNessuna valutazione finora
- Respuesta Inmune en InvertebradosDocumento18 pagineRespuesta Inmune en InvertebradosWendyVanessaPulgarinOvalleNessuna valutazione finora
- Inmunidad innata y adaptativa: definición y componentesDocumento2 pagineInmunidad innata y adaptativa: definición y componentesevil25Nessuna valutazione finora
- Cabello blanco en alopecia areata: formas clínicas y mecanismos propuestosDocumento23 pagineCabello blanco en alopecia areata: formas clínicas y mecanismos propuestosFabian BedoutNessuna valutazione finora
- Catalogo Pe PDFDocumento33 pagineCatalogo Pe PDFjheysonNessuna valutazione finora
- Validación de Pruebas Serológicas para El Diagnóstico de Enfermedades InfecciosasDocumento20 pagineValidación de Pruebas Serológicas para El Diagnóstico de Enfermedades InfecciosasmagnerNessuna valutazione finora
- PlaneaciónDocumento19 paginePlaneaciónnidia iselaNessuna valutazione finora
- Emociones y Sistema Inmune, PPT Autor: Dr. Orestes García FigueiredoDocumento61 pagineEmociones y Sistema Inmune, PPT Autor: Dr. Orestes García FigueiredoNancySáezNessuna valutazione finora
- Linfoma de HodgkinDocumento27 pagineLinfoma de HodgkinAlexis GomezNessuna valutazione finora
- Sarcoidosis, HX, LLM, Microlitiasis, NEUMOSUR PDFDocumento14 pagineSarcoidosis, HX, LLM, Microlitiasis, NEUMOSUR PDFJosé PérezNessuna valutazione finora
- Monocitopoyesis: Formación de monocitos y macrófagosDocumento19 pagineMonocitopoyesis: Formación de monocitos y macrófagosMariana TurizoNessuna valutazione finora
- BADILLO MARTINEZ ISAAC - Docx Procedimientos para La Protección Del HospederoDocumento2 pagineBADILLO MARTINEZ ISAAC - Docx Procedimientos para La Protección Del HospederoLupizz BlueeNessuna valutazione finora
- Conamad Copeerativa MadridDocumento40 pagineConamad Copeerativa MadridMaricuelina Omega TresNessuna valutazione finora
- TAREA 2 CuestionarioDocumento3 pagineTAREA 2 CuestionarioBerenize VidalNessuna valutazione finora
- Neisseria GonorrhoeaeDocumento6 pagineNeisseria Gonorrhoeaesarai villenaNessuna valutazione finora
- Una Vacuna Es Una Suspensión de Microorganismos Vivos AtenuadosDocumento4 pagineUna Vacuna Es Una Suspensión de Microorganismos Vivos AtenuadosLina PérezNessuna valutazione finora
- Actividad 7 Sistema Nervioso y Sistema InmunologicoDocumento8 pagineActividad 7 Sistema Nervioso y Sistema InmunologicoMerlinaNessuna valutazione finora
- Enfermedades reproductivas, terneros y planes sanitarios bovinosDocumento80 pagineEnfermedades reproductivas, terneros y planes sanitarios bovinosRuben Danilo Rondon Pereira0% (1)
- Método de Burri para diagnóstico de CryptococosisDocumento9 pagineMétodo de Burri para diagnóstico de CryptococosisMaria Cruz BlassNessuna valutazione finora
- Biologia NavarraDocumento33 pagineBiologia NavarraHelena Ojuel GómezNessuna valutazione finora
- INFORME S2 ESO26 Bioquímica Seminario - Copia FaltDocumento7 pagineINFORME S2 ESO26 Bioquímica Seminario - Copia FaltCoraine Chávez CórdovaNessuna valutazione finora
- Final IiDocumento6 pagineFinal IiLetícia Raquel PintoNessuna valutazione finora
- AnafilaxiaDocumento33 pagineAnafilaxiaNicolas De la CruzNessuna valutazione finora
- Proyecto Hillary Jahely Intriago MeroDocumento28 pagineProyecto Hillary Jahely Intriago MeroBenigna PonceNessuna valutazione finora
- INMUNOGENETICADocumento78 pagineINMUNOGENETICAMiguel VasquezNessuna valutazione finora
- Vasos e Ganglios Linfaticos FinalDocumento8 pagineVasos e Ganglios Linfaticos FinalAlane OliveiraNessuna valutazione finora
- Tres Clases.Documento111 pagineTres Clases.Alejandro BenitesNessuna valutazione finora
- Morfologicas y Definicion de Los LeucocitosDocumento13 pagineMorfologicas y Definicion de Los LeucocitosJorge Vidal100% (1)