Potrebbero piacerti anche
- Cómo Alargar El Pene Naturalmente en Casa Con EjerciciosDocumento24 pagineCómo Alargar El Pene Naturalmente en Casa Con Ejerciciosdaniellhe charles100% (1)
- Costas Orlando Comunicacion Por Medio de La Predicacion PDFDocumento239 pagineCostas Orlando Comunicacion Por Medio de La Predicacion PDFAlejandro Morales Ruiz100% (3)
- Codigos de Falla ChryslerDocumento151 pagineCodigos de Falla ChryslerRobert Nunez84% (25)
- Vendaje Neuromuscular Kinesiology Tape ManualDocumento70 pagineVendaje Neuromuscular Kinesiology Tape Manualchedeybaez100% (3)
- Ayudas ergogénicas y nutricionalesDa EverandAyudas ergogénicas y nutricionalesValutazione: 5 su 5 stelle5/5 (2)
- Desconexion ParticularDocumento1 paginaDesconexion ParticularRonald Monzón100% (1)
- Cuestionario DASHDocumento6 pagineCuestionario DASHPaula Ortiz OyarzunNessuna valutazione finora
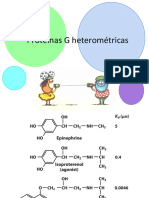
- Enfermedades Hereditarias Que Implican Proteínas GDocumento7 pagineEnfermedades Hereditarias Que Implican Proteínas GUSMP FN ARCHIVOS67% (3)
- Teoria Glutamatergica de La EsquizofreniaDocumento6 pagineTeoria Glutamatergica de La EsquizofreniaEstefanía Valencia HenaoNessuna valutazione finora
- Receptores Acoplados A ProteinasDocumento7 pagineReceptores Acoplados A Proteinaspain667Nessuna valutazione finora
- Proteina GDocumento6 pagineProteina GAngela Gonzales HerreraNessuna valutazione finora
- 2RECEP18 Protei-Nas G y Sistema AdenililciclasaDocumento15 pagine2RECEP18 Protei-Nas G y Sistema AdenililciclasaHbk Denver HendersonNessuna valutazione finora
- Receptores de MembranaDocumento9 pagineReceptores de MembranaAlexy SanchezNessuna valutazione finora
- Acetilcolina en Muscula Cardiaco PDFDocumento10 pagineAcetilcolina en Muscula Cardiaco PDFGonzalo UribeNessuna valutazione finora
- Bases Estructurales de Los ReceptoresDocumento12 pagineBases Estructurales de Los ReceptoresSalvador LauraNessuna valutazione finora
- Examen FisiologíaDocumento34 pagineExamen FisiologíaAna Elia Hernández GonzálezNessuna valutazione finora
- Receptores, Segundos Mensajeros y Vias de Senalizacion1Documento14 pagineReceptores, Segundos Mensajeros y Vias de Senalizacion1SantiagoNessuna valutazione finora
- Proteina GDocumento6 pagineProteina GThalina PayaresNessuna valutazione finora
- Apuntes Proteinas GDocumento40 pagineApuntes Proteinas GYulsAelbert ReRoNessuna valutazione finora
- 5. Receptores, Segundos Mensajeros y Vias de Senalizacion1Documento14 pagine5. Receptores, Segundos Mensajeros y Vias de Senalizacion1esrn4laboratorioNessuna valutazione finora
- Proteinas GDocumento3 pagineProteinas Gcorrales.gersonNessuna valutazione finora
- Proteínas G: estructura, clasificación, funciones y su importancia en la transducción de señalesDocumento19 pagineProteínas G: estructura, clasificación, funciones y su importancia en la transducción de señalesEduardo P. FernándezNessuna valutazione finora
- Seminario #8 (Ii-3) Semana #11 Desordenes en Receptores CelularesDocumento15 pagineSeminario #8 (Ii-3) Semana #11 Desordenes en Receptores CelularesSantiago Delgado ChumioqueNessuna valutazione finora
- Recepción Celular. Mecanismos MolecularesDocumento17 pagineRecepción Celular. Mecanismos MolecularesTEFANessuna valutazione finora
- Microclases ReceptoresDocumento5 pagineMicroclases ReceptoresMilagros BritesNessuna valutazione finora
- Los Receptores Que Son Acoplados A Proteínas GDocumento12 pagineLos Receptores Que Son Acoplados A Proteínas GAlberto SilvaNessuna valutazione finora
- de Proteína GDocumento10 paginede Proteína GEvelin Alexandra Orozco MurgasNessuna valutazione finora
- GLUCOCORTICOIDESDocumento15 pagineGLUCOCORTICOIDESDANIEL ALEJANDRONessuna valutazione finora
- AMPLIAR EL ESPECTRO DE LAS ENFERMEDADES Proteína GDocumento13 pagineAMPLIAR EL ESPECTRO DE LAS ENFERMEDADES Proteína GFranco RomeroNessuna valutazione finora
- Prot G Adenilato CiclasaDocumento5 pagineProt G Adenilato CiclasaVictor VitoriaNessuna valutazione finora
- Hormona GHDocumento4 pagineHormona GHbelencecyte38Nessuna valutazione finora
- 6.1 Cuales Son Los Mediadores Por Segundos Mensajeros Que Intervienen en Las Respuestas Celulares A Las Hormonas Proteicas o PeptídicasDocumento1 pagina6.1 Cuales Son Los Mediadores Por Segundos Mensajeros Que Intervienen en Las Respuestas Celulares A Las Hormonas Proteicas o PeptídicasNicole LoraNessuna valutazione finora
- Taller de Señalización CelularDocumento6 pagineTaller de Señalización CelularKevin PalacioNessuna valutazione finora
- Receptores HormonalesDocumento4 pagineReceptores Hormonalessebastian OrbeaNessuna valutazione finora
- Señalización IntracelularDocumento3 pagineSeñalización IntracelularALONDRA MONTSERRAT GRADILLA SILVANessuna valutazione finora
- GTPDocumento2 pagineGTPPaul VargasNessuna valutazione finora
- RECEPTORES ACOPLADOS A LA PROTEINA G Practica IDocumento5 pagineRECEPTORES ACOPLADOS A LA PROTEINA G Practica IJose CarlosNessuna valutazione finora
- 6-Hormonas y Mecanismo de AccionDocumento4 pagine6-Hormonas y Mecanismo de AccionJuan Fuentes100% (1)
- De Receptores Transmembrana: Rutas ProteolíticasDocumento125 pagineDe Receptores Transmembrana: Rutas ProteolíticasPablo SPNessuna valutazione finora
- Karp BiologiaCelular SEÑALIZACIONDocumento47 pagineKarp BiologiaCelular SEÑALIZACIONInazuma EstradaNessuna valutazione finora
- TRADUCCIONDocumento2 pagineTRADUCCIONAngie Nathalia MartinezNessuna valutazione finora
- Via de Las Pentosas y GlutationDocumento16 pagineVia de Las Pentosas y GlutationAlexander Salinas CalderónNessuna valutazione finora
- SeñalizacionDocumento4 pagineSeñalizacionFranciscoNessuna valutazione finora
- Señalización Celular, Camilo Escobedo Fernández PDFDocumento5 pagineSeñalización Celular, Camilo Escobedo Fernández PDFcamiloesferNessuna valutazione finora
- Qué es una hormonaDocumento10 pagineQué es una hormonaEdgardoNessuna valutazione finora
- Proteinas G ArticulosDocumento15 pagineProteinas G ArticulosElizabethNessuna valutazione finora
- Mecanismo de Acción de La ProlactinaDocumento3 pagineMecanismo de Acción de La ProlactinaMichael SaavedraNessuna valutazione finora
- Qué Es Una Proteína GDocumento1 paginaQué Es Una Proteína GErick Ramos OlivaNessuna valutazione finora
- GTPDocumento4 pagineGTPANGEL C.Nessuna valutazione finora
- PREGUNTAS BIOLOGÍA CELULAR KarpDocumento13 paginePREGUNTAS BIOLOGÍA CELULAR KarpEduardo Lara Magaña100% (2)
- Resume Del Cap de BioquimicaDocumento3 pagineResume Del Cap de Bioquimicanayeliceli2345Nessuna valutazione finora
- Mecanismo de Acción de Los ReceptoresDocumento9 pagineMecanismo de Acción de Los ReceptoresPaola LaraNessuna valutazione finora
- Glucagon y Transduccion de SeñalesDocumento2 pagineGlucagon y Transduccion de SeñalesJohn Curtis Castrejón50% (2)
- Las Proteínas G HeterotriméricasDocumento28 pagineLas Proteínas G HeterotriméricasAbigail IturraldeNessuna valutazione finora
- MECANISMOS DE ACCIÓN Parte 2Documento7 pagineMECANISMOS DE ACCIÓN Parte 2MARIA ALEJANDRA BERMUDEZ BUITRAGONessuna valutazione finora
- Fisio. Integracion EndocrinaDocumento11 pagineFisio. Integracion EndocrinaGeranni RondonNessuna valutazione finora
- GMPCDocumento8 pagineGMPCRominaRojasSoto0% (1)
- Aprende las tres cascadas de señalización clave: AMPc, insulina y fosfoinositolDocumento3 pagineAprende las tres cascadas de señalización clave: AMPc, insulina y fosfoinositolMilton Alexi Sepulveda Castro100% (1)
- AMPc y El GMPCDocumento9 pagineAMPc y El GMPCVivi EstefaniaNessuna valutazione finora
- Bioquimica Ii Vias de Transducción de Señales: Dra. (C) Emma Urrunaga Soria Profesora Del CursoDocumento61 pagineBioquimica Ii Vias de Transducción de Señales: Dra. (C) Emma Urrunaga Soria Profesora Del CursoClinton LunaNessuna valutazione finora
- GTPASADocumento5 pagineGTPASAANGEL C.Nessuna valutazione finora
- Metabolismo Del GlucógenoDocumento2 pagineMetabolismo Del GlucógenoLuana HuertasNessuna valutazione finora
- Proteinas GDocumento19 pagineProteinas GAnonymous 443xsmeqtNessuna valutazione finora
- Taller Biología CelularDocumento4 pagineTaller Biología CelularMarianaNessuna valutazione finora
- ApendicitisDocumento6 pagineApendicitisDanna cárdenasNessuna valutazione finora
- 554-Texto Del Artículo-3564-1-10-20200728Documento20 pagine554-Texto Del Artículo-3564-1-10-20200728Ešmęrąldą ŤôrôNessuna valutazione finora
- 1.1) Ruidos Cardiacos, Semiología de SoplosDocumento8 pagine1.1) Ruidos Cardiacos, Semiología de SoplosDanna cárdenasNessuna valutazione finora
- BALANTIDIUM COLI UltimoDocumento9 pagineBALANTIDIUM COLI UltimoDanna cárdenasNessuna valutazione finora
- VPH y cáncer de cérvix: causas, tipos de VPH, epidemiología y patogeniaDocumento12 pagineVPH y cáncer de cérvix: causas, tipos de VPH, epidemiología y patogeniaDanna cárdenasNessuna valutazione finora
- Análisis Terapia de La Hormona Del Crecimiento en Los NiñosDocumento4 pagineAnálisis Terapia de La Hormona Del Crecimiento en Los NiñosDanna cárdenasNessuna valutazione finora
- 1) Semiología Torácica y PulmónDocumento9 pagine1) Semiología Torácica y PulmónDanna cárdenasNessuna valutazione finora
- 2.2) FeocromositomaDocumento5 pagine2.2) FeocromositomaDanna cárdenasNessuna valutazione finora
- Analisis de La Obra Los Miserables - Victor HugoDocumento10 pagineAnalisis de La Obra Los Miserables - Victor HugoDanna cárdenasNessuna valutazione finora
- Arco Reflejo 1ra y 2da MotoneuronaDocumento5 pagineArco Reflejo 1ra y 2da MotoneuronaMelissa SuarezNessuna valutazione finora
- 2) AMIBIASIS y GIARDIASISDocumento5 pagine2) AMIBIASIS y GIARDIASISDanna cárdenasNessuna valutazione finora
- Analisis de Articulo L.CDocumento16 pagineAnalisis de Articulo L.CDanna cárdenasNessuna valutazione finora
- GlucolisisDocumento5 pagineGlucolisisDanna cárdenasNessuna valutazione finora
- Punto 1Documento1 paginaPunto 1Danna cárdenasNessuna valutazione finora
- Sintomas y EstadisticasDocumento1 paginaSintomas y EstadisticasDanna cárdenasNessuna valutazione finora
- Via SorbitolDocumento4 pagineVia SorbitolDanna cárdenasNessuna valutazione finora
- CK-BB: Tejido Cerebral Y Pulmonar: Debido A Que Lack-Bb Se Encuentra enDocumento3 pagineCK-BB: Tejido Cerebral Y Pulmonar: Debido A Que Lack-Bb Se Encuentra enDanna cárdenasNessuna valutazione finora
- Producción Del Óxido Nítrico DoDocumento24 pagineProducción Del Óxido Nítrico DoDanna cárdenasNessuna valutazione finora
- Paralisis Tercer Par CranealDocumento4 pagineParalisis Tercer Par CranealDanna cárdenas100% (1)
- TransaminasasDocumento4 pagineTransaminasasDanna cárdenasNessuna valutazione finora
- Nuevo Codigo Codigo de Transito Ley 1383 de 2010 PDFDocumento30 pagineNuevo Codigo Codigo de Transito Ley 1383 de 2010 PDFRuben HuffNessuna valutazione finora
- Receptores Del Cuerpo HumanoDocumento1 paginaReceptores Del Cuerpo HumanoDanna cárdenasNessuna valutazione finora
- HallazgosDocumento10 pagineHallazgosOscar MGNessuna valutazione finora
- INTRODUCCIÓN E HISTORIA WolfDocumento1 paginaINTRODUCCIÓN E HISTORIA WolfDanna cárdenasNessuna valutazione finora
- Mi Ángel C2 2024-1Documento192 pagineMi Ángel C2 2024-1ordonezmagdalena75Nessuna valutazione finora
- Certificado Estudios SecundariaDocumento2 pagineCertificado Estudios SecundariaJhon Laura BenavidesNessuna valutazione finora
- Quimica IsomeríaDocumento9 pagineQuimica IsomeríaJohan Manuel Gamez MendozaNessuna valutazione finora
- Regularización migratoria decreto supremoDocumento2 pagineRegularización migratoria decreto supremodavid galvisNessuna valutazione finora
- Diseño y Simulación de Sistemas de Manufactura - Foro 2-01.08.2021Documento2 pagineDiseño y Simulación de Sistemas de Manufactura - Foro 2-01.08.2021janett ramirezNessuna valutazione finora
- Sesión 7-8 - Gestión de Ventas Prom 45Documento55 pagineSesión 7-8 - Gestión de Ventas Prom 45Gustavo SalvatierraNessuna valutazione finora
- Examen Matemáticas III 2da VueltaDocumento3 pagineExamen Matemáticas III 2da VueltaWenzel Dietrich ValentinaNessuna valutazione finora
- Aisladores SismicosDocumento11 pagineAisladores SismicosYadira VelasquezNessuna valutazione finora
- Implantación de La Estrategia LogísticaDocumento45 pagineImplantación de La Estrategia LogísticaAbel Insua LagoNessuna valutazione finora
- 1 y 2. Concepto e Import An CIA Del SueloDocumento55 pagine1 y 2. Concepto e Import An CIA Del Sueloands_120Nessuna valutazione finora
- Tarea VirtualDocumento6 pagineTarea VirtualRyan J Salas VeraNessuna valutazione finora
- Bajo Piso Tekno Step PDFDocumento3 pagineBajo Piso Tekno Step PDFRicci RománNessuna valutazione finora
- Juegos Con Nombres de BebesDocumento2 pagineJuegos Con Nombres de BebesOmar GonzálezNessuna valutazione finora
- Practica No 1 - Generador de Señal de RelojDocumento6 paginePractica No 1 - Generador de Señal de RelojAngel JaraNessuna valutazione finora
- Ca6 Unidad Didactica Sistemas ContabesDocumento85 pagineCa6 Unidad Didactica Sistemas ContabesAndreina Macias MendozaNessuna valutazione finora
- Promesa Parques de La Bocha C-201 PrivadaDocumento3 paginePromesa Parques de La Bocha C-201 PrivadaJORGE H. VARELANessuna valutazione finora
- Internet Cables SubmarinosDocumento2 pagineInternet Cables SubmarinosDolores Cristóbal María SofíaNessuna valutazione finora
- Proyecto Tipo PPBC Derecho San MarcosDocumento3 pagineProyecto Tipo PPBC Derecho San MarcosJuan RobertoNessuna valutazione finora
- Cartilla Co-Cre-Ar 2016 PDFDocumento84 pagineCartilla Co-Cre-Ar 2016 PDFdanilo rosasNessuna valutazione finora
- Caso Clinico ABP - Grupo 3 - Estructura y Funcion 1Documento5 pagineCaso Clinico ABP - Grupo 3 - Estructura y Funcion 1ZuffelyEstelaRojasNessuna valutazione finora
- Taller Fundamentos ElectricosDocumento4 pagineTaller Fundamentos Electricosjose estebanNessuna valutazione finora
- Alisva Cárdenas-Pérez: Agradecimiento ESPE LatacungaDocumento1 paginaAlisva Cárdenas-Pérez: Agradecimiento ESPE LatacungaWalter DiablithoNessuna valutazione finora
- Cambio de La Demanda y Cantidad DemandadaDocumento3 pagineCambio de La Demanda y Cantidad DemandadaLeo FernandezNessuna valutazione finora
- PORTAFOLIO DE TRABAJOS DE ARQUITECTURA 2013, Arq Oscar Daniel Contreras CaroDocumento31 paginePORTAFOLIO DE TRABAJOS DE ARQUITECTURA 2013, Arq Oscar Daniel Contreras Carooscar daniel contrerasNessuna valutazione finora