Potrebbero piacerti anche
- Temas poderosos e reflexões sobre nome e destinoDocumento45 pagineTemas poderosos e reflexões sobre nome e destinoFérnanda Moura100% (4)
- Ossos do membro torácico em equinosDocumento8 pagineOssos do membro torácico em equinosMarielle LimaNessuna valutazione finora
- Livro de OrtopticaDocumento67 pagineLivro de OrtopticaPaula EmyNessuna valutazione finora
- Aula Parametros Genéticos PDFDocumento20 pagineAula Parametros Genéticos PDFKaio FernandesNessuna valutazione finora
- Melhoramento Animal: herdabilidade e correlação genéticaDocumento10 pagineMelhoramento Animal: herdabilidade e correlação genéticaRosely Mattos De O QuintanilhaNessuna valutazione finora
- MRP SAP Estudo conceitosDocumento18 pagineMRP SAP Estudo conceitosTuaregue_BRNessuna valutazione finora
- Manual de Confecção de Resenhas EquinasDocumento24 pagineManual de Confecção de Resenhas EquinasJosiane MacielNessuna valutazione finora
- Dimensionamento de rosca sem fim e coroa para motor de 6 CVDocumento7 pagineDimensionamento de rosca sem fim e coroa para motor de 6 CVWilsonNessuna valutazione finora
- Osteologia Prática - 01Documento10 pagineOsteologia Prática - 01Ester Beghini0% (1)
- Atlas de osteologia bovinos equinosDocumento21 pagineAtlas de osteologia bovinos equinosJoão Victor Lopes BastosNessuna valutazione finora
- Geometria Sagrada - Robert LlawlorDocumento116 pagineGeometria Sagrada - Robert Llawlorzmercedez96% (24)
- Melhoramento Genetico em Gado LeiteiroDocumento41 pagineMelhoramento Genetico em Gado LeiteiroDelano AlencarNessuna valutazione finora
- Laboratório 1 - Cruzamentos e Estatística AnimalDocumento3 pagineLaboratório 1 - Cruzamentos e Estatística AnimalAmanda TramonteNessuna valutazione finora
- 5 - Herdabilidade e RepetibilidadeDocumento47 pagine5 - Herdabilidade e RepetibilidadeBrunoMuchaleNessuna valutazione finora
- Propriedade ou pessoa?: os animais na sociedade e na Medicina Veterinária: uma proposta abolicionistaDa EverandPropriedade ou pessoa?: os animais na sociedade e na Medicina Veterinária: uma proposta abolicionistaNessuna valutazione finora
- Estação de MontaDocumento24 pagineEstação de MontaJoao Paulo TeixeiraNessuna valutazione finora
- Atividades de GramáticaDocumento33 pagineAtividades de GramáticaDebora RibeiroNessuna valutazione finora
- Manejo reprodutivo de gado de corteDocumento71 pagineManejo reprodutivo de gado de corteMirian FernandesNessuna valutazione finora
- Ossos do membro anteriorDocumento5 pagineOssos do membro anteriorTamylin NagasawaNessuna valutazione finora
- Miologia VeterináriaDocumento4 pagineMiologia VeterináriaCibele De Sousa Vanderley CostaNessuna valutazione finora
- Bovinos de Corte CruzamentosDocumento83 pagineBovinos de Corte Cruzamentosa097792Nessuna valutazione finora
- Nutrição de búfalas leiteiras: composição, requerimentos e formulação de raçõesDocumento16 pagineNutrição de búfalas leiteiras: composição, requerimentos e formulação de raçõesJadir FélixNessuna valutazione finora
- Estudo tempo instalação alterações cadavéricas cãesDocumento6 pagineEstudo tempo instalação alterações cadavéricas cãesTalita D'Paula100% (1)
- Funcao Quadratica ProblemasDocumento3 pagineFuncao Quadratica ProblemasSolange PalmeiraNessuna valutazione finora
- Fisiologia da digestão em ruminantesDocumento70 pagineFisiologia da digestão em ruminantesAmanda Hellen Silva de AndradeNessuna valutazione finora
- Patologia Clínica CirúrgicaDocumento17 paginePatologia Clínica CirúrgicadaniellesilvavvieiraNessuna valutazione finora
- Secreções digestivas e regulaçãoDocumento48 pagineSecreções digestivas e regulaçãoBetânia Glória CamposNessuna valutazione finora
- Indicadores do estresse térmico em bovinosDocumento11 pagineIndicadores do estresse térmico em bovinossufealNessuna valutazione finora
- 3.endocrinologia e Puberdade No MachoDocumento55 pagine3.endocrinologia e Puberdade No MachoCelly Sousa100% (2)
- Leis de Newton exercíciosDocumento11 pagineLeis de Newton exercíciosIsabelle LopesNessuna valutazione finora
- Determinação Da Idade de Bovinos Atraves Dos Dentes ProntoDocumento16 pagineDeterminação Da Idade de Bovinos Atraves Dos Dentes ProntoJoana Alves100% (2)
- Estratégia Cruzamento (Bov. Leiteiros)Documento18 pagineEstratégia Cruzamento (Bov. Leiteiros)Tassiê TurcatoNessuna valutazione finora
- Aula 7 - NNP e Aditivos para RuminantesDocumento58 pagineAula 7 - NNP e Aditivos para Ruminanteslucas marquesNessuna valutazione finora
- Sistema Locomotor de Equinos - Aula 12Documento58 pagineSistema Locomotor de Equinos - Aula 12Maria Isabel Alves BarbosaNessuna valutazione finora
- Zootecnia: Davi Matos Emile Lorhane Eloah Lima Evilly BritoDocumento12 pagineZootecnia: Davi Matos Emile Lorhane Eloah Lima Evilly BritoPedro VitorNessuna valutazione finora
- Exame Ginecológico em Pequeno AnimaisDocumento7 pagineExame Ginecológico em Pequeno AnimaisGrazi HonorioNessuna valutazione finora
- Avaliação Trimestral - Matemática - 4.º AnoDocumento7 pagineAvaliação Trimestral - Matemática - 4.º AnoCarla PereiraNessuna valutazione finora
- Introduã A Anatomia VeterinãriaDocumento42 pagineIntroduã A Anatomia VeterinãriaLarissa Gobbi100% (2)
- Cinética espermática e fertilidade: avaliação de parâmetrosDocumento41 pagineCinética espermática e fertilidade: avaliação de parâmetrosLet_cia_Morais_6660Nessuna valutazione finora
- Genética quantitativa: herança Mendeliana x poligênicaDocumento75 pagineGenética quantitativa: herança Mendeliana x poligênicaDiogo JuniorNessuna valutazione finora
- Afecções da Glândula Mamária em RuminantesDocumento27 pagineAfecções da Glândula Mamária em RuminantesAmanda AliceNessuna valutazione finora
- Sistema reprodutor feminino em animaisDocumento11 pagineSistema reprodutor feminino em animaisIsabelle PassosNessuna valutazione finora
- Aprumos de Equinos e BovinosDocumento21 pagineAprumos de Equinos e BovinosPaulo Eduardo Ferreira dos Santos paulo.eduardoNessuna valutazione finora
- 2 Lista de Exercicios VetDocumento3 pagine2 Lista de Exercicios Vetallan felipeNessuna valutazione finora
- Estudo Dirigido Anatomia Veterinária I EsqueletoDocumento4 pagineEstudo Dirigido Anatomia Veterinária I EsqueletoClésia BarretoNessuna valutazione finora
- Babesia: Parasita Hemoparasita Transmitido por CarrapatosDocumento28 pagineBabesia: Parasita Hemoparasita Transmitido por CarrapatosGarcía Méndez MelissaNessuna valutazione finora
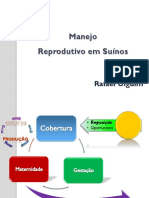
- Manejo Reprodutivo em SuínosDocumento55 pagineManejo Reprodutivo em SuínosPedro SantosNessuna valutazione finora
- Apostila - Anato I Muito BoaDocumento62 pagineApostila - Anato I Muito BoaDiego MunizNessuna valutazione finora
- Aula 2 - Genética de Populações PDFDocumento54 pagineAula 2 - Genética de Populações PDFClaudemir TrevisanNessuna valutazione finora
- 1 - Fisiologia Do Sistema NervosoDocumento67 pagine1 - Fisiologia Do Sistema NervosoDébora BandeiraNessuna valutazione finora
- Envenenamento Equino Pela Casca de Café (CoffeaDocumento20 pagineEnvenenamento Equino Pela Casca de Café (CoffeaJuliana Barroso Félix100% (1)
- Sexagem EspermáticaDocumento16 pagineSexagem EspermáticaALICE CAROLINANessuna valutazione finora
- Genética PopulaçõesDocumento47 pagineGenética PopulaçõesAmanda CristinaNessuna valutazione finora
- Cronologia Dentaria Dos BovinosDocumento4 pagineCronologia Dentaria Dos BovinosJana Kelly S. SeiNessuna valutazione finora
- Roteiro de aulas práticas de histologia da Universidade Federal da Grande DouradosDocumento66 pagineRoteiro de aulas práticas de histologia da Universidade Federal da Grande DouradosKenia GomezNessuna valutazione finora
- Osteologia de MembrosDocumento16 pagineOsteologia de MembroskaliusiaNessuna valutazione finora
- Atualidades e Tendências Da EquinoculturaDocumento14 pagineAtualidades e Tendências Da EquinoculturakcauabreuNessuna valutazione finora
- Anestesia na cesariana de cadelasDocumento35 pagineAnestesia na cesariana de cadelasSidnei H. Vieira JúniorNessuna valutazione finora
- 03 Ossos Do CrânioDocumento31 pagine03 Ossos Do CrânioCesar Henrique LinderNessuna valutazione finora
- Implementação da identificação equina em PortugalDocumento23 pagineImplementação da identificação equina em PortugalFátima RibeiroNessuna valutazione finora
- Osteologia Axial Cabeça ZOOTEC 2014Documento93 pagineOsteologia Axial Cabeça ZOOTEC 2014Fábio FrançaNessuna valutazione finora
- Anatomia e Fisiologia Das AvesDocumento42 pagineAnatomia e Fisiologia Das AvesWilson SilvaNessuna valutazione finora
- Músculos e suas funçõesDocumento34 pagineMúsculos e suas funçõesThais Nunes100% (1)
- Resolução Cirúrgica de Cólicas em EquinosDocumento88 pagineResolução Cirúrgica de Cólicas em EquinosPaulo Y. ImaiNessuna valutazione finora
- Avaliação Diagnóstica sobre Raças de BovinosDocumento9 pagineAvaliação Diagnóstica sobre Raças de BovinosDeise VieiraNessuna valutazione finora
- Alguns Pontos Importantes em Exames Clínicos de AnimaisDocumento4 pagineAlguns Pontos Importantes em Exames Clínicos de AnimaisVet_arquivosNessuna valutazione finora
- AviculturaDocumento128 pagineAviculturaDenise Cristina de OliveiraNessuna valutazione finora
- Pelagem de EquinosDocumento18 paginePelagem de EquinosSamara SouzaNessuna valutazione finora
- Onde está o Gato de SchrödingerDocumento2 pagineOnde está o Gato de SchrödingerbarsumgmailNessuna valutazione finora
- Manual de Estudos de Tráfego DNIT 2006Documento388 pagineManual de Estudos de Tráfego DNIT 2006dougx20Nessuna valutazione finora
- Apostila HP12C BarretoDocumento70 pagineApostila HP12C BarretoHenilton SantosNessuna valutazione finora
- Aula 15 Ciclo de Roscamento - TorneamentoDocumento6 pagineAula 15 Ciclo de Roscamento - TorneamentoJunior GuedesNessuna valutazione finora
- Sistema de bombeamento de águaDocumento9 pagineSistema de bombeamento de águaTarcisio Kock FilhoNessuna valutazione finora
- Calculo - Rosca TransportadoraDocumento4 pagineCalculo - Rosca TransportadoraEder MendesNessuna valutazione finora
- Exercicios para Fixação - Lógica de ProgramaçãoDocumento4 pagineExercicios para Fixação - Lógica de ProgramaçãoIgor Maurício SantosNessuna valutazione finora
- FRP Análise Numérica e ExperimentalDocumento9 pagineFRP Análise Numérica e ExperimentalJose Eduardo GranatoNessuna valutazione finora
- Aula 03-Distribuicoes - de - ProbabilidadeDocumento78 pagineAula 03-Distribuicoes - de - ProbabilidadenakyoNessuna valutazione finora
- Treino Matemático 8o anoDocumento4 pagineTreino Matemático 8o anoAna Carla de OliveiraNessuna valutazione finora
- Revisão de TrigonometriaDocumento39 pagineRevisão de TrigonometriaAlex MarquesNessuna valutazione finora
- Prova PB Matematica 4ano Tarde 4bim PDFDocumento6 pagineProva PB Matematica 4ano Tarde 4bim PDFTamika OwensNessuna valutazione finora
- Função Do Primeiro Grau E Suas Aplicações: Function of The First Degree and Its ApplicationsDocumento9 pagineFunção Do Primeiro Grau E Suas Aplicações: Function of The First Degree and Its ApplicationsRebecca heloise Oliveira ValérioNessuna valutazione finora
- ML8000DDocumento42 pagineML8000Ddanielwjr0% (2)
- Betão pré-esforçado - Verificação ELU flexão e cortanteDocumento20 pagineBetão pré-esforçado - Verificação ELU flexão e cortanteMacdonald NdjiveNessuna valutazione finora
- NBR 5382 - Verificação de Iluminação de Interiores PDFDocumento4 pagineNBR 5382 - Verificação de Iluminação de Interiores PDFMateus DauricioNessuna valutazione finora
- p16 MatematicaDocumento20 paginep16 MatematicadanielifrnNessuna valutazione finora
- ListarotacoesDocumento7 pagineListarotacoesdnielemilaioNessuna valutazione finora
- Ef11 Teste Avaliacao 1 CCDocumento3 pagineEf11 Teste Avaliacao 1 CCLaura MonteiroNessuna valutazione finora
- Trabalho de EstaleiroDocumento10 pagineTrabalho de EstaleiroMarijó NeuriceNessuna valutazione finora