Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
Resumen Langman Apuntes Completo
Caricato da
L JCopyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
Formati disponibili
Resumen Langman Apuntes Completo
Caricato da
L JCopyright:
Formati disponibili
lOMoARcPSD|4000730
Resumen langman - Apuntes completo
Histología y Embriología (Universidad Nacional de La Plata)
StuDocu no está patrocinado ni avalado por ningún colegio o universidad.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
LANGMAN’i
Embriología Médica
GAMETOGÉNESIS: transformación de las células germinales en gametos femeninos y masculinos
CÉLULAS GERMINALES PRIMIGENIAS
El desarrollo comienza con la fecundación (fusión de los gametos masculino y femenino). Los gametos derivan de las células
germinales primigenias (CGP) que se forman en el epiblasto durante la 2ª semana y posteriormente se trasladan a la pared del saco
vitelino. Durante la 4ª semana, estas células empiezan a migrar desde el saco vitelino hasta las gónadas en desarrollo, donde llegan
hacia el final de la 5ª semana. El nro. de divisiones aumenta durante la migración y cuando las células ya han alcanzado las gónadas.
Para prepararse p/la fecundación, las células germinales experimentan el proceso de gametogénesis, que incluye una meiosis,
p/reducir el nro. de cromosomas, y el de citodiferenciación p/acabar de madurar.
CAMBIOS MORFOLÓGICOS DURANTE LA MADURACIÓN DE LOS GAMETOS
Ovogénesis
Es el proceso mediante el cual los ovogonios se diferencian en ovocitos maduros.
La maduración de los ovocitos se inicia antes del nacimiento
Una vez que las CGP han alcanzado la gónada de una mujer, se diferencian en ovogonios. Estas células experimentan
diversas divisiones mitóticas y, hacia el final del 3er mes, se disponen en grupos rodeados por una capa de céls epiteliales planas.
Mientras que es probable que todos los ovogonios de un grupo procedan de una misma célula, las céls epiteliales planas=céls
foliculares, se originan a partir del epitelio superficial que recubre el ovario.
La mayoría de los ovogonios continúan dividiéndose por mitosis, pero algunos de ellos detienen sus divisiones celulares en la
profase de la meiosis I y forman ovocitos primarios. Durante los meses siguientes, el nro. de ovogonios aumenta rápidamente y hacia
el 5º mes de desarrollo prenatal el nro. total de céls germinales en el ovario alcanza su cifra máxima, estimada en 7 millones. En este
momento las céls empiezan a morir y muchos ovogonios y ovocitos primarios degeneran y se vuelven atrésicos. Hacia el 7º mes la
mayoría de los ovogonios han degenerado, excepto unos cuantos que se encuentran en la superficie. Todos los ovocitos primarios
supervivientes han entrado en la profase de la meiosis I y la mayoría están rodeados por una capa individual de céls foliculares
epiteliales planas. El conjunto formado por un ovocito primario y las céls epiteliales planas que le rodean se conoce como folículo
primordial
La maduración de los ovocitos continúa en la pubertad
Cuando se acerca el momento del nacimiento, todos los ovocitos han iniciado la profase de la meiosis I, pero en lugar de
continuar en metafase, continúa en la fase de diploteno, una etapa de reposo durante la profase que se caracteriza por una red laxa de
cromatina. Los ovocitos primarios se detienen en la profase y no completarán su primera división meiotica hasta la pubertad. Esta fase
de reposo es inducido por el inhibidor de la maduración del ovocito (IMO), un pequeño péptido que segregan las células foliculares.
Se estima que en el momento del nacimiento el nro. total de ovocitos varía de 600mil a 800mil. Durante la infancia, la maduración de
ovocitos se vuelven atrésicos; al inicio de la pubertad solo quedan unos 40mil, de los cuales se ovularan menos de 500. Algunos de los
ovocitos que alcanzan la madurez en las etapas tardías de la vida antes de ser ovulados permanecen inactivos en la fase de diploteno
de la meiosis I durante 40años o más.
En la pubertad se establece una reserva de folículos en crecimiento que se mantiene gracias al suministro de folículos
primordiales. Cada mes, entre 15 a 20 folículos de esta reserva empiezan a madurar, y en el proceso pasan por tres etapas: 1) primaria
o prenatal (ovocito primario), 2) secundaria o antral (ovocito secundario o vesicular) y 3) preovulatoria (folículo de de Graaf)
En cada ciclo ovárico empiezan a desarrollarse unos cuantos folículos, pero generalmente solo uno alcanza la madurez. Los
otros degeneran y se vuelven atrésicos. Cuando el folículo secundario ha fecundado, una descarga de LH induce la fase de crecimiento
preovulatoria. Se completa la meiosis I, lo que lleva a la formación de dos céls hijas de tamaño desigual, cada una con 23 cromosomas
dobles. Una de estas células, el ovocito secundario, recibe la mayor parte del citoplasma; la otra, el primer corpúsculo polar,
prácticamente no recibe citoplasma. El primer corpúsculo polar se dispone entre la zona pelúcida y la membrana celular del ovocito
secundario en el espacio perivitelino. A continuación, la célula entra en la meiosis II, pero se detiene en la metafase aproximadamente
3h antes de la ovulación. La meiosis II solo se completa si el ovocito es fecundado; en caso contrario la célula degenera
aproximadamente 24h después de la ovulación. El primer corpúsculo polar puede experimentar una segunda división.
Espermatogénesis
La maduración de los espermatoioides se inicia en la pubertad
La espermatogénesis incluye todos aquellos acontecimientos por los cuales los espermatogonios se transforman en
espermatozoides.
En el momento del nacimiento, en los cordones testiculares de un varón RN pueden reconocerse las céls germinales rodeadas
por céls de sostén. Las céls de sostén derivan del epitelio superficial de la glándula, se convierten en céls sustentaculares o céls de
Sertoli.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
Poco antes de la pubertad, los cordones espermáticos adquieren una luz y se convierten en túbulos seminíferos.
Aproximadamente al mismo tiempo, las CGP originan céls precursoras de espermatogonios. A intervalos regulares, emergen céls de
esta población de céls madres q forman espermatogonios te tipo A espermatogonios de tipo B espermatocitos primarios. En
este momento, los espermatocitos primarios entran en una profase larga (22días) y seguidamente completan con rapidez la meiosis I y
forman los espermatocitos secundarios. Durante la meiosis II, estas céls empiezan a formar espermátidas. A lo largo de esta serie de
acontecimientos, la citocinesis permanece incompleta, de manera q las sucesivas generaciones de céls quedan unidas por puentes
citoplasmáticos. Además, mientras se desarrollan, los espermatogonios y las espermátidas permanecen dentro de cavidades profundas
de céls de Sertoli.
La espermatogénesis está regulada por la LH. LH=céls de Leydig testosterona=céls de Sertoli espematogénesis. La
FSH tb es esencial, se une a las céls de Sertoli
Espermiogénesis
Es la serie de cambios que tansforman las espermátidas en espermatozoides. Estos cambios consisten en 1) formación del
acrosoma q cubre la 1/2del núcleo, 2)la condensación del núcleo, 3) la formación del cuello, la pieza intermedia y la cola, 3) el
desprendimiento de la mayor parte del citoplasma en forma de cuerpos residuales
El tiempo que requiere un espermatogonio p/convertirse en espermatozoide maduro es aproximadamente de 74 días, y cada
día se producen 300millones de espermatozoides.
Cuando están completamente formados los espermatozoides túbulos seminíferos epidídimo (donde adquieren la
movilidad completa)
PRIMERA SEMANA DEL DESARROLLO: de la ovulación a la implantación
Ciclo ovárico
En la pubertad, la mujer empieza a experimentar ciclos menstruales regulares. La FSH y la LH estimulan y controlan los
cambios cíclicos en el ovario.
Al inicio de cada ciclo ovárico, la FSH estimula e/15 a 20 folículos en fase primaria (prenatal) p/q crezcan. En condiciones
normales solo uno de estos folículos alcanzará la plena madurez, y únicamente se liberará un ovocito; los otros folículos degeneran y
se vuelven atrésicos. La FSH tb estimula la maduración de las céls foliculares (granulosa) q rodean el ovocito. A su vez la
proliferación de estas céls está rodeada por el factor 9 de la transformación del crecimiento β (TGF- β).las células de la tec interna
producen testoterona en respuesta a LH. Las céls de la teca int y las céls granulosas cooperan p/producir estrógenos. Debido a la
producción de estrógenos:
• El endometrio uterino entra en la fase folicular o proliferativa
• El moco cervical se adelgaza p/permitir el paso del esperma, y
• Se estimula la adenohipófisis p/q produzca LH
A ½ del ciclo se produce una descarga de LH que:
• Eleva la concentración del factor promotor de la maduración, lo q induce los ovocitos a
completar la meiosis I e iniciar la meiosis II
• Estimula la producción de progesterona q parte de las céls del estroma foliculares
(luteinización), y
• Provoca la ruptura del folículo y la ovulación
Ovulación
Durante los días inmediatamente después de la ovulación, bajo influencia de LH y FSH, el folículo 2rio crece con rapidez
hasta alcanzar un diámetro de 25mm. Al mismo tiempo un aumento brusco de LH induce el ovocito 1rio a completar la meiosis I y
hace q el folículo entre en la fase preovulatoria. Asimismo se inicia la meiosis II, pero el ovocito se detiene en la metafase
aproximadamente 3h antes de la ovulación
Cuerpo lúteo
Después de la ovulación, bajo la influencia de la LH, las céls de la granulosa y de la teca int remanentes forman el cuerpo
lúteo y secretan estrógenos y progesterona. Esta última, junto con parte del estrógeno, hace que la mucosa uterina entre en la fase
progestacional o secretora y se prepare p/la implantación del embrión.
Transporte del ovocito
La acción de barrido de las fimbrias de la trompas hace caer el ovocito en la trompa de Falopio. La velocidad de transporte
está regulada por el estado endocrino durante y después de la ovulación. El ovocito fecundado llega a la luz del útero al cabo de
aproximadamente 3 ó 4 días.
Cuerpo albicans
Si no hay fecundación, el cuerpo lúteo alcanza su máximo desarrollo a los 9 días después de la ovulación. Más tarde el
cuerpo lúteo degenera y forma una masa de tejido fibroso cicatrizante=cuerpo albicans. Al mismo tiempo, la producción de
progesterona disminuye y precipita el sangrado menstrual.
Si el ovocito es fecundado la GCh secretada por los sincitiotrofoblastos evita la degeneración del cuerpo lúteo. Este continúa
creciendo y forma el cuerpo lúteo del embarazo. Al término del 3er mes, el tamaño de esta estructura equivale a 1/3 o ½ del tamaño
del ovario. Las céls amarillas continúan secretando progesterona hasta el final del 4ºmes; a partir de entonces, se retiran lentamente
hasta q la función de estas sea reemplazada por la placenta.
Fecundación
Es el proceso mediante el cual los gametos fem y masc se fusionan, esto tiene lugar en la ampolla de la trompa uterina (a 2/3
del útero). Los espermatozoides pueden mantenerse viables en el tracto reproductor de la mujer durante varios días (48 a 72 h)
Solo el 1% de los espermatozoides depositados en la vagina entran en el cuello del útero, donde sobreviven unas cuantas h.
El viaje de los espermatozoides desde el cuello del útero hasta la trompa de Falopio requiere un mínimo de 2h y 7h.
P/q los espermatozoides puedan fecundar el ovocito, deben experimentar un proceso de: 1) capacitación (aprox 7 h), durante
la cual pierden la cubierta glucoproteica y las proteínas plasmáticas seminales de su cabeza, y 2) una reacción acrosómica, durante la
cual se liberan sustancias del tipo de la tripsina y la acrosina p/poder penetrar la zona pelúcida. Durante la fecundación, el
espermatozoide debe penetrar en 1) la corona radiada, 2) la iona pelúcida, y 3) la membrana celular del ovocito.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
En el momento en que el espermatozoide ha penetrado el ovocito: 1) el ovocito termina su meiosis II y forma el pronúcleo
femenino, 2) la zona pelúcida se vuelve impenetrable p/otros espermatozoides, y 3) la cabeza del espermatozoide se desprende de la
cola, se engruesa y forma el pronúcleo masculino. Una vez que el ADN de ambos núcleos se ha replicado, los cromosomas materno y
paterno se entremezclan, se dividen longitudinalmente y experimentan una división mitótica, lo que resulta en una fase bicelular.
c La fecundación da lugar a: 1) restablecimiento del nro. diploide de cromosomas, 2) determinación del sexo cromosómico, y
3) inicio de la segmentación
Segmentación
Una vez que el cigoto ha llegado a la fase de dos céls, experimenta una serie de divisiones mitóticas que aumentan el nro. de
células. Estas, cuyo tamaño se reduce en cada división, reciben el nombre de blastómeros. Hasta la fase de ocho céls, forman un grupo
laxo. Después de la 3ª división los blastómeros maximizan el contacto e/ellos y forman una pelota compacta de céls q se mantienen
juntas con uniones herméticas. Este proceso de compactación separa las céls internas, q están totalmente comunicadas mediantes
uniones intercelulares comunicantes, de las céls externas.
Aproximadamente 3días después de la fecundación, las céls del embrión compactado se dividen de nuevo y forman una
mórula de 16 céls (mora). Las céls internas de la mórula constituyen la masa celular interna (origina los tejidos propios del embrión)
y las céls que la rodean forman la masa celular externa (forma el trofoblasto, que más adelante contribuirá a la formación de la
placenta)
La fase de dos céls se alcanza aproximadamente 30 h después de la fecundación; la de 4 céls se logra hacia las 40 h, la de 12
a 16 céls hacia los 3días, y la fase final de la mórula, aprox a los 4 días. La zona pelúcida desaparece al final del 4º día
Formación de los blastocitos
Aproximadamente en el momento que la mórula entra en la cavidad uterina, a través de la zona pelúcida empieza a penetrar
líquido dentro de los espacios intercelulares de la masa celular interna. De forma gradual estos espacios confluyen y acaban formando
una cavidad única=el blastocele. En este momento el embrión es un blastocito.
Las céls de la masa celular interna, ahora llamada embrioblasto, se encuentra en un polo, mientras que las céls de la masa
celular externa o trofoblasto, se aplanan y forman la pared epitelial del blastocito. La zona pelúcida ha desaparecido, lo que posibilita
la implantación. Las céls trofoblásticas del polo del embrioblasto empiezan a penetrar e/las céls de la mucosa uterina hacia el 6º día.
Por lo tanto, al final de la 1ª semana del desarrollo, el cigoto humano ha pasado por los estados de mórula y blastocito y ha
empezado a implantarse en la mucosa uterina
El útero en el momento de la implantación
En el momento de la implantación, el útero se encuentra en la fase secretora (se inicia 2 ó 3 días después de la ovulación) y el
blastocito se implanta en el endometrio de la pared anterior o posterior. Si no hay fecundación, se inicia la fase menstrual y se
desprenden los estratos esponjoso y compacto del endometrio. El estrato basal del endometrio se conserva para regenerar los otros
estratos durante el siguiente ciclo.
SEGUNDA SEMANA DEL DESARROLLO: el disco bilaminar
• Cap 4: Segunda semana de desarrollo
• Dia 8: blastocito casi sumergido en endometrio. Los trofoblastos se diferencia en
citotrofoblastos (capa interna mononucleada con act mitotica) y sincitiotrofoblastos
(multinucleadas). Las cels de la masa interna se diferencia en dos capas: hipoblastica (cels
cubicas adyacentes a cavidad de blastocito) y epiblastica ( cels cilíndricas adyacentes a
cav amniótica). Ambas capas forman un disco plano con una cavidad interna que se
convierte en cavidad amniótica. Las cels epiblasticas adyacentes a los citotrofoblastos se
denominan amnioblastos (revisten cav amniótica junto con epiblasto)
• Dia 9: blastocito inmerso en endometrio. Aparecen vacuolas a nivel del sincitio
endometrial que se fusionan entre ellas formando lagunas (periodo lagunar). Una capa de
cels del hipoblasto forman memb exocelomica de Heuser que reviste sup interna de
citotrofoblasto formando cav exocelomica junto con el hipoblasto.
• Dia 11/12: se forma un bulto en pared uterina. Se forman los sinusoides de las lagunas
que dan capilares revestidos por citotrofoblastos. Entre cels de la capa externa de cav
exocelomica y cels internas de citotrofoblasto se forma una cavidad llamada celoma
extraembionario o cavidad corionica. Capa que reviste amnios y citotrofoblasto:
mesodermo somatopleurico extraembrionario: Capa que reviste saco vitelino: mesodermo
esplacnopleurico extraembrionario.
• Dia 13: Puede ocurrir sangrado uterino (confunde con mestruación). Proliferan cels de
citotrofoblasto formando vellosidades primarias. Cels de hipoblasto proliferan revistiendo
sacovitelino primitivo, formando saco vitelino secundario definitivo. Pueden desprenderse
segmentos de cav exocelomica (saco vitelino 1rio) que forman quistes exocelomicos. El
celoma extraembrionario se agranda formando cav corionica revestida por placa corionica
(mesodermo extraembrionario).
•
• Cap 5 Tercera semana: disco trilaminar
• Se caracteriza por GASTRULACIÓN (formacionde disco trilaminar: ectodermo,mesodermo,
endodermo). Se inicia con formación de línea primitiva en superficie de epiblasto. En el
extremo cefálico de la línea se forma el nodulo primitivo que roda a la fosita primitiva. Las
cels del epiblasto migran hacia esta línea y la invaginan (gracias a factor de crecimiento
fibroblastico 8). Luego de invaginarse, desplazan al hipoblasto formando endodermo,
otras se situan en el medio formando endodermo, y el resto forma ectodermo. Las cels se
expanden a los bordes y en polo cefálico forman placa precordal.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
• Formacion de notocorda: las cels prenotocordales del nodulo primitivo migran a placa
precordal formando dos capas que constituten la placa notocordal, cuyas cels proliferan
luego formando la notocorda definitiva que sirve de base al esqueleto axial (se encuentra
inf a tubo neural). La pared post del saco vitelino forma un divertículo (alantoides) en el
16to dia que en vertebrados inferiores sirve como deposito de desechos renales.
• Establecimiento de ejes corporales: el endodermo visceral anterior expresa genes
esenciales p/formación de cabeza (OTX3,L1M1,HESX1,cerberus,lefty). En presencia de prot
morfogenica osea 4 y factor de crecimiento de fibroblastos, el mesodermo se desplaza
ventralmente p/formar rinhones, sangre y mesodermo de la pared lateral del cuerpo. En
presencia del factor de transcr. Goosecoid el mesodermo se desplaza caudalmente.
Nogina y folistina son antagonistas también de prot morfogenica osea 4 (BMP4). Nodal
interviene en formación/mantenimiento de línea primitiva. HNF3beta mantiene nodulo
primitvo (sin este no hay gastrulación->no hay prosecefalo y mesencéfalo). Nodal
+Lefty2=regulan PITX2 que establece lateralidad izquierda. Sonic hedgehog evita que
estos genes actúen a la derecha. Se cree que gen Snail regula lateralidad derecha.
• El disco embrinario crece caudalemente de forma principal. La migración de cels a la línea
primitiva se mantiene hasta semana 4.
• Desarrollo trofoblastico: Al iniciarse 3ra semana, se forman vellosidade primarias.
Luego las cels mesodérmicas penetran es esta formando las vellosidades secundarias. Al
final de la 3ra semana las cels de las vellosidades secundarias empiezan a diferenciarse
formando vellosidades terciarias o placentarias definitivas que están en contacto con
capilares del pediculo. Las cels del citotrofoblasto penetran en endometrio y forman
cubierta citotrofoblastica externa. Esta cubierta va rodeando al trofoblasto
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
• PERIODO FETAL (desde el 3º mes hasta la fecha de parto)
Desarrollo del feto
En general se considera que la duración de la gestación es de 280 días o 40 semanas, pero de
manera más exacta 266 días o 38 semanas después de la fecundación.
El periodo fetal se caracteriza por la maduración de los tejidos y órganos y por el crecimiento del
cuerpo. Durante este periodo se producen muy pocas malformaciones.
3º, 4º y 5º mes: mayor incremento de longitud (se expresa en vértice – nalga, VN, o vértice – talón,
VT. La longitud aumenta 5 cm. por mes).
2 últimos meses: mayor incremento de peso (700 gr. por mes).
Cambios según los meses
El desarrollo de la cabeza se vuelve más lento en comparación con el resto del cuerpo:
3º mes: ½ longitud.
5º mes: 1/3 longitud.
Nacimiento: ¼ longitud.
3º mes
Los ojos se localizan en la superficie ventral de la cara, las orejas se sitúan cerca de su
posición definitiva.
Las extremidades alcanzan su longitud relativa con respecto al resto del cuerpo (siendo las
inferiores más cortas y menos desarrolladas que las superiores).
Se encuentran los centros de osificación primaria en huesos largos y del cráneo.
Los genitales externos se han desarrollado lo suficiente como para ser determinados por
medio de exámenes.
Las asas intestinales se retraen del cordón umbilical hacia la cavidad abdominal.
4º y 5º mes en adelante
El feto aumenta de longitud rápidamente (al termino del 6º mes su longitud V-N es de
aproximadamente 15 cm. – la mitad de la longitud del recién nacido-.).
El peso aumenta poco, al final de 5º mes no alcanza los 500 gr.
El feto esta cubierto por un vello delicado – lanugo-. Son visibles el cabello y las cejas.
Durante el 5º mes los movimientos del feto son percibidos por la madre.
Durante el 6º mes el peso comienza a aumentar notablemente. El feto parece arrugado por la
falta de tejido conectivo subyacente y su piel es rojiza.
En los 2 últimos meses se redondea el contorno corporal como consecuencia del depósito de
grasa subcutánea. Hacia el final de la vida intrauterina la piel esta cubierta por una sustancia grasosa
blanquecina – vernix casesoa -, producto de secreción de las glándulas sebáceas.
Al final del 9º mes el cráneo tiene mayor circunferencia que cualquier otra parte del cuerpo.
En la fecha de nacimiento el peso del feto es de 3000 a 3400 gr., su longitud V-N es de 36 cm. y
la longitud V-T es de 50 cm. aproximadamente.
Aumento de peso y longitud durante el periodo fetal
EDAD LONGITUD V-N
PESO (gr.)
(semanas) (cm.)
9-12 5-8 10-45
13-16 9-14 60-200
17-20 15-19 250-450
21-24 20-23 500-820
25-28 24-27 900-1300
29-32 28-30 1400-2100
33-36 31-34 2200-2900
37-38 35-36 3000-3400
MEMBRANAS FETALES Y PLACENTA
En el 2º mes el trofoblasto se caracteriza por poseer abundantes vellosidades secundarias y
terciarias (ancladas en el mesodermo de la lamina corionica y unidas a la decidua materna por medio
de la envoltura citotrofobalstica externa) que le dan un aspecto radiado. La superficie de las
vellosidades esta formada por el sincitio que descansa sobre células citotrofoblasticas que cubren la
parte central del mesodermo vascularizado. El sistema capilar que se desarrolla en el centro de las
vellosidades se pone en contacto con los capilares de la lamina corionica y del pedículo de fijación
originando el sistema vascular extraembrionario.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
En los meses siguientes, de los troncos de las vellosidades salen abundantes prolongaciones
que se dirigen hacia los espacios intervellosos o lacunares. Estas vellosidades pierden las células
trofoblasticas y algunas células del tejido conectivo hacia el 4º mes. Entonces las únicas capas que
separan la circulación materna y la fetal son el sincitio y la pared endotelial de los vasos sanguíneos.
La desaparición de las células citotrofoblasticas avanza desde las vellosidades menores hasta
las mayores, aunque siempre persisten algunas en las vellosidades más grandes, sin embargo, estas
no participan en el intercambio entre las dos circulaciones.
Corion frondoso y decidua basal
En las primeras semanas de desarrollo las vellosidades cubren toda la superficie del corion. A
medida que avanza la gestación las vellosidades del polo embrionario se dilatan y forman el corion
frondoso, las del polo abembrionario o vegetativo degeneran hacia el 3º mes formando una porción
lisa, el corion leve o calvo.
Otra diferencia entre los polos embrionario y abembrionario es la decidua (capa funcional del
endometrio, se desprende durante el parto). La decidua basal, que cubre al corion frondoso, consiste
en una capa de células deciduales voluminosas con abundantes lípidos y glúcogeno. La decidua
capsular cubre al polo abembrionario o vegetativo y degenera con el aumento de volumen de la
vesícula corionica. En una etapa posterior, el corion leve se pone en contacto con la pared uterina
(decidua parietal) en el lado opuesto del útero y las dos capas se fusionan obliterando la cavidad
uterina.
La única porción del corion que participa de los procesos de intercambio es el corion frondoso
que forma con la decidua basal la placenta.
La fusión del amnios y el corion forma la membrana amniocoriónica que oblitera la cavidad
corionica y que se rompe al iniciar el trabajo de parto.
Estructura de la placenta
Hacia el comienzo del 4º mes la placenta posee dos porciones:
Porción materna constituida por la decidua basal.
Porción fetal constituida por el corion frondoso.
En el lado fetal la placenta esta rodeada por la lamina corionica. En el lado materno, la lámina
decidual de la decidua basal se incorpora íntimamente a la placenta. En la zona de unión se
entremezclan células gigantes del trofoblasto (sincitiales) y las deciduales. Esta zona posee un
abundante material extracelular amorfo. En este momento la mayor parte de las células
citotrofobasticas han degenerado.
Entre la lamina corionica y la decidual se encuentran los espacios intervellosos ocupados por
sangre materna, que provienen de las lagunas sincitiotrofobasticas revestidos por sincitio de origen
fetal. Las vellosidades arborescentes se desarrollan en los lagos sanguíneos intervellosos.
En el curso de 4º y 5º mes la decidua forma los tabiques deciduales que sobresalen en los
espacios intervellosos pero no llegan a la lamina corionica. Estos tabiques poseen un núcleo central de
tejido materno, pero su superficie esta cubierta por una capa de células sincitiales que separa la
sangre materna del tejido fetal de las vellosidades. La formación de los tabiques divide la placenta en
compartimientos o cotiledones manteniéndose el contacto entre los espacios intervellosos.
La placenta aumenta de tamaño durante la gestación y cubre aproximadamente entre el 15 y 30%
de la superficie interna del útero. El aumento de grosor de la placenta se debe a la arborización de las
vellosidades existentes.
Placenta de término
Es discoide, tiene un diámetro de 15 a 25 cm. aproximadamente, 3 cm. de espesor y pesa
alrededor de 500 a 600 gr.
En el momento del parto se desprende de la pared uterina y unos 30 minutos después del parto
es expulsada de la cavidad del útero.
El lado materno posee 15 a 20 cotiledones (zonas sobresalientes) cubiertos por una delgada
capa de decidua basal y separados por los tabiques deciduales.
El lado fetal esta cubierto completamente por la lamina corionica donde se observan arterias y
venas de grueso calibre – vasos coriónicos – que convergen en el cordón umbilical. A su vez el corion
esta cubierto por el amnios.
Circulación de la placenta
Los cotiledones reciben sangre a través de las arterias espirales que en numero de 80 a 100
atraviesan la lamina decidual y entran en los espacios intervellosos.
La luz de la arteria espiral es reducida, lo cual produce un aumento de la presión de la sangre
al entrar al espacio intervelloso donde baña a las vellosidades arborizadas con sangre oxigenada. Al
disminuir la presión la sangre fluye de retorno desde la lamina corionica hacia la decidua, donde entra
en las venas endometriales (drenando así hacia la circulación materna).
Los espacios intervellosos contienen aproximadamente 150 ml de sangre que se recambia 3 a 4 veces
por minuto y se desplazan a lo largo de las vellosidades corionicas que tienen un área superficial que
varia entre 4 a 14 m2.
Este recambio no tiene lugar en todas las vellosidades, sino solo en las que están en íntimo
contacto con la membrana sincitial. En estas vellosidades el sincitio posee un ribete en cepillo formado
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
por numerosas microvellosidades que aumentan el índice de recambio entre la circulación materna y
la fetal.
La membrana placentaria separa la sangre materna de la fetal y esta formada por 4 capas en un
principio:
El revestimiento endotelial de los vasos fetales
El tejido conectivo del núcleo de las vellosidades
La capa citotrofoblastica
El sincitio
A partir del 4º mes en adelante, la membrana placentaria se adelgaza ya que el revestimiento
endotelial se pone en íntimo contacto con la membrana sincitial aumentando el índice de recambio.
Se considera que la placenta es de tipo hemocoriónico, ya que separa la sangre materna de la fetal
mediante un derivado corionico.
Funciones de la placenta
Intercambio de gases: el intercambio de oxígeno, dióxido de carbono y monóxido de carbono
se realiza mediante difusión simple. El feto extrae 20 a 30 ml de oxigeno por minuto de la
circulación materna (la cantidad de oxigeno depende de la oferta y no de la difusión).
Intercambio de elementos nutritivos y de electrolitos: como aminoácidos, ácidos grasos
libres, hidratos de carbono y vitaminas.
Transmisión de anticuerpos maternos: los anticuerpos son captados por pinocitosis por el
sincitiotrofoblasto y llevados después a los capilares fetales. La inmunoglobulina G (IgG)
recibida de la madre protege al feto de diversas enfermedades infecciosas adquiriendo
inmunidad pasiva contra la difteria, viruela, sarampión, pero no así contra la varicela.
Producción de hormonas: hacia el final del 4º mes la placenta produce progesterona como
para mantener la gestación en caso de extirpación o falla del cuerpo amarillo. Produce también
hormonas estrógenas (estradiol) hasta antes de completarse la gestación, donde alcanza su
nivel máximo contribuyendo al desarrollo de la gandula mamaria y al crecimiento del útero,
gonadotrofinas (gonadotrofina corionica humana – hCG) que tienen un efecto parecido al de las
hormonas luteinizantes y son excretadas por la madre en la orina (indicador de embarazo),
somatomamotrofina (antes llamada lactógeno placentario) sustancia semejante a la hormona
del crecimiento que le confiere al feto prioridad sobre la glucosa sanguínea materna.
Amnios y cordón umbilical
La línea de reflexión entre el amnios y el ectodermo embrionario es ovalada y se denomina
anillo umbilical primitivo. En la quinta semana de desarrollo pasan por esta estructura: el pedículo
de fijación (incluye la alantoides y los vasos umbilicales – 2 arterias y 1 vena), el pedículo del saco
vitelino (conducto onfalomesenterico) acompañado por los vasos vitelinos y el conducto que comunica
las cavidades celómicas extra e intraembrionarias.
El saco vitelino propiamente dicho ocupa un espacio en cavidad corionica (espacio entre el
amnios y la lamina corionica).
La cavidad amniótica crece a expensas de la cavidad corionica y el amnios comienza a envolver
a los pedículos de fijación y al saco vitelino formando el cordón umbilical primitivo.
En sentido distal el cordón comprende el pedículo del saco vitelino y los vasos umbilicales. En
sentido proximal incluye algunas asas intestinales y el resto de la alantoides.
El saco vitelino se encuentra en la cavidad corionica unido al cordón umbilical por su pedículo.
Hacia el final del 3º mes el amnios se ha dilatado y se pone en contacto con el corion
obliterando la cavidad corionica.
Algunas asas intestinales sobresalen hacia el celoma extraembrionario en el cordón umbilical,
formando la hernia umbilical fisiológica. Hacia el final del 3º mes las asas vuelven al cuerpo del
embrión y desaparece la cavidad celomica en el cordón umbilical. Cuando se obliteran además la
alantoides, el conducto vitelino y los vasos, solo quedan en el cordón los vasos umbilicales rodeados
por la gelatina de Wharton (tejido rico en proteoglucanos que funciona como capa protectora). En el
feto de termino el cordón umbilical tiene 2 cm. de diámetro y 50 a 60 cm. de longitud, es tortuoso y
presenta nudos falsos.
Cambios de la placenta hacia el final del embarazo
Aumento del tejido fibroso en el centro de las vellosidades.
Mayor grosor de la membrana basal de los capilares fetales.
Cambios de obliteración de los capilares de pequeño calibre de las vellosidades.
Deposito de sustancia fibrinoide en la superficie de la vellosidades de la zona de union y en la
lamina corionica.
Liquido amniótico
Liquido acuoso y cristalino producido por parte de las células amnióticas, pero que proviene
principalmente de la sangre materna y ocupa la cavidad amniótica.
La cantidad aumenta desde 30 ml a las 10 semanas hasta 350 ml a las 20 semanas y de 800 a 1000
ml a las 37 semanas. El liquido:
Amortigua las sacudidas.
Impide que el embrión se adhiera al amnios.
Permite los movimientos fetales.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
El volumen de líquido es reemplazado cada 3 horas. A partir del 5º mes el feto ingiere
aproximadamente 400 ml del líquido amniótico por día (50% del volumen total). Durante este mes
también se añade orina (en su mayor parte es agua, ya que la placenta actúa en intercambio de los
desechos metabólicos) diariamente al liquido amniótico
Membranas fetales en gemelos
Gemelos dicigóticos, biovulares o fraternos
Resultan de la expulsión simultánea de 2 ovocitos y de la fecundación por 2 espermatozoides
diferentes. Ambos cigotos se implantan en forma individual en el útero y casa uno de ellos desarrolla
su placenta, su amnios y su saco corionico. A veces las 2 placentas están muy juntas y pueden
fusionarse, lo mismo ocurre con el saco amniótico. En ocasiones los gemelos dicigóticos poseen
eritrocitos de dos tipos diferentes – mosaicismo eritrocítico -.
Gemelos monocigóticos, uniovulares o idénticos
Se desarrollan a partir de un solo ovulo fecundado. Son el resultado de que el cigoto se divida en
diferentes etapas de desarrollo. Se considera que la separación comienza en el periodo bicelular, en
cuyo caso se desarrollan 2 cigotos diferentes. Ambos blastocistos se implantan independientemente y
cada embrión posee su propia placenta y saco corionico. La separacion del cigoto tiene lugar en el
periodo del blastocisto, cuando la masa celular interna se divide en 2 grupos de células dentro de la
misma cavidad del blastocisto y los 2 embriones tienen en común la placenta y la cavidad corionica,
pero cavidades amnióticas diferentes. Cuando la separación se produce en el periodo del disco
germinativo bilaminar se forman 2 embriones que comparten una misma placenta, sacos amnióticos y
coriónicos.
SISTEMA NERVIOSO CENTRAL
Aparece al inicio de la 3º semana como una placa alargada de ectodermo engrosado – placa
neural -, situada en la región dorsal media por delante de la fosita primitiva. Poco después sus bordes
se elevan formando los pliegues neurales. Los pliegues neurales se acercan en la línea media, se
une y forman el tubo neural. La fusión comienza en la región cervical, continuando en dirección
cefálica y caudal donde se retarda y temporalmente los neuroporos cefálico y caudal comunican la luz
del tubo neural con la cavidad amniótica. El cierre del neuroporo cefálico tiene lugar en el periodo de
18 a 20 somitas (25º día); el neuroporo caudal se oblitera unos días mas tarde.
El extremo cefálico presenta 3 dilataciones: las vesículas encefálicas primarias
(prosencéfalo o cerebro anterior, mesencéfalo o cerebro medio y rombencéfalo o cerebro posterior).
Simultáneamente se forman 2 plegamientos: el pliegue cervical (en la unión del rombencéfalo y la
medula espinal) y el pliegue cefálico (en la región del mesencéfalo).
Cuando el embrión tiene 5 semanas, el prosencéfalo esta formado por 2 porciones: el
telencéfalo o cerebro terminal constituido por una parte media y dos evaginaciones laterales –
hemisferios cerebrales – y el diencéfalo que se caracteriza por la evaginacion de las vesículas ópticas.
El mesencéfalo esta separado del rombencéfalo por un surco profundo – surco de His -. El
rombencéfalo esta compuesto por 2 porciones: el metencéfalo (que formará la protuberancia y el
cerebelo) y el mielencéfalo, separados por el pliegue protuberencial.
La luz de la medula espinal – conducto ependimario - se continúa con la cavidad de las
vesículas encefálicas.
La cavidad del rombencéfalo es el cuarto ventrículo, la del diencéfalo es el tercer ventrículo y la
de los hemisferios cerebrales son los ventrículos laterales.
El tercer y cuarto ventrículo se comunican a través de la luz del mesencéfalo – acueducto de
Silvio -. Los ventrículos laterales se comunican con el tercer ventrículo a través del foramen de Monro.
Médula espinal
CAPAS NEUROEPITELIAL, DEL MANTO Y MARGINAL
La pared del tubo neural esta formada por células neuroepiteliales que forman un grueso
epitelio pseudoestratificado y están conectadas por complejos de unión que se encuentran en la luz.
Después de cerrarse el tubo, estas células se dividen rápidamente y producen mayor cantidad de
células neuroepiteliales que en conjunto forman el neuroepitelio.
Una vez que se ha cerrado el tubo, las células neuroepiteliales originan a los neuroblastos o células
nerviosas primitivas (con un gran núcleo redondo que posee un nucleoplasma pálido y un nucleolo que
se tiñe de color oscuro) que rodean a la capa neuroepitelial y se denomina capa de manto que formará
la sustancia gris de la medula espinal.
La capa mas externa de la medula espinal contiene las fibras nerviosas que salen de los
neuroblastos y se denomina capa marginal.
Como consecuencia de la mielinización de las fibras nerviosas, esta capa adquiere un aspecto blanco
por lo que se llama sustancia blanca de la medula espinal.
PLACAS BASALES, ALARES, DEL TECHO Y DEL PISO
La continua adición de neuroblastos a la capa del manto origina 2 engrosamientos a cado lado
del tubo neural, uno ventral y otro dorsal. Los engrosamientos ventrales o placas basales incluyen a
las células motoras de las astas ventrales y forman las áreas motoras de la medula. Los
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
engrosamientos dorsales o placas alares forman las áreas sensitivas. El surco limitante señala el
límite entre las placas. Las porciones dorsal (placa de techo) y ventral (placa de piso) no poseen
neuroblastos y sirven como vías para las fibras nerviosas que cruzan de un lado a otro.
Además de las astas motora ventral y sensitiva dorsal, entre las dos áreas se acumula un grupo
de neuronas que formara el asta lateral, que contiene neuronas del sistema nervioso autónomo y solo
se encuentra a nivel torácico y lumbar superior (L2-L3).
DIFERENCIACION HISTOLOGICA
Células nerviosas
Los neuroblastos se originan exclusivamente por división de las células neuroepiteliales. En su
primer momento tienen una prolongación central que se extiende hacia la luz (dendrita pasajera) que
desaparece al emigrar hacia la zona del manto con lo que adoptan una forma redonda temporal –
neuroblastos apolares -. Luego aparecen 2 nuevas prolongaciones citoplasmáticas a los lados del
cuerpo celular formando un neuroblasto bipolar. La prolongación de un extremo de la célula se alarga
rápidamente para formar el axón primitivo o cilindroeje, en tanto que el otro extremo presenta
arborizaciones citoplasmáticas que constituyen las dendritas primitivas. En esta etapa la célula se
denomina neuroblasto multipolar y con el desarrollo ulterior se convierte en neurona. Una vez
formados, los neuroblastos pierden la capacidad de dividirse. Los cilindroejes de las neuronas en la
placa basal atraviesan la zona marginal y se manifiestan en la zona ventral de la medula espinal (raíz
motora ventral del nervio raquídeo o espinal, conducen los impulsos motores de la medula hacia los
músculos). Los cilindroejes de las neuronas de la placa alar penetran en la capa marginal donde
ascienden o descienden a otros niveles para formar neuronas de asociación.
Células de la glía
Los glioblastos son formados por las células neuroepiteliales cuando ha cesado la producción de
neuroblastos. Emigran desde la capa neuroepitelial hacia la capa del manto y la marginal. En la capa
del manto se diferencian en astrocitos protoplasmáticos y astrocitos fibrosos.
La célula de la oligodendroglia deriva de los glioblastos y se encuentra principalmente en la
capa marginal formando las vainas de mielina que rodean a los axones.
En la segunda mitad de la vida intrauterina aparecen las células de la microglia, derivadas del
mesenquima y de tipo fagocítico.
Cuando las células neuroepiteliales dejan de producir neuroblastos y glioblastos se diferencian a
células ependimarias que tapizan el conducto central de la medula espinal.
Células de la cresta neural
Aparecen durante el plegamiento de la placa neural a cada lado del surco neural. Son de origen
ectodérmico y forman temporalmente una zona intermedia entre el ectodermo y el tubo neural desde
el prosencéfalo hasta el nivel de los somitas caudales y lateralmente hasta la cara dorsolateral del
tubo neural. Algunas células originan los ganglios sensitivos de los nervios raquídeos.
Los neuroblastos presentan 2 prolongaciones:
Centrípetas: penetran en la porción dorsal del tubo neural, en la medula espinal terminan en el
asta dorsal o ascienden por la capa marginal hasta los centros cerebrales superiores. Forman la
raíz sensitiva dorsal del nervio raquídeo.
Centrifugas: se unen a las fibras de la raíz motora ventral contribuyendo a la formación del
nervio raquídeo.
Las células de la cresta neural también se diferencian a neuroblastos simpáticos, células de
Scwhann, células pigmentarias, odontoblastos, meninges y mesenquima de los arcos faringeos.
Nervios espinales o raquídeos
Las fibras nerviosas motoras comienzan a aparecer en la 4º semana de desarrollo intrauterino y
se originan en células nerviosas de la placas basales, reuniéndose en haces para formar las raíces
nerviosas ventrales.
Las raíces nerviosas dorsales se originan en ganglios de la raíz dorsal (ganglios espinales). Las
prolongaciones centrales de estos ganglios forman haces que se desarrollan en el lado opuesto a las
astas dorsales. Las prolongaciones dístales se unen a raíces ventrales para formar un nervio espinal o
raquídeo que se divide en ramos primarios dorsales (musculatura axial dorsal, articulaciones
intervertebrales y piel de la espalda) y ramos primarios ventrales (miembros y la pared ventral.
Forman los principales plexos neviosos).
Mielinización
La mielinización de los nervios periféricos es llevada a cabo por las células de Scwhann (se
originan en la cresta neural) que forman la vaina de Scwhann o neurilema a partir del 4º mes.
La vaina de mielina que rodea a las fibras nerviosas de la medula espinal es formada por
células de la oligodendroglia aproximadamente al 4º mes de vida intrauterina y se extiende hasta el 1º
año de vida postnatal.
MODIFICACIONES DE LA POSICION DE LA MEDULA ESPINAL
En el 3º mes de desarrollo la medula se extiende en toda la longitud del embrión y los nervios
raquídeos atraviesan los agujeros intervertebrales en su nivel de origen.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
Al nacimiento el extremo se encuentra a nivel de la 3º lumbar y los nervios raquídeos tienen una
dirección oblicua desde su segmento de origen en la medula espinal hasta el nivel correspondiente en
la columna.
En el adulto termina a la altura de L2-L3, formando por debajo el filum terminale (prolongación
de piamadre) que señala el camino de retroceso de la medula y se encuentra unido a la primera
vértebra coccígea. Las fibras nerviosas que se encuentran por debajo del extremo terminal de la
medula forman la cauda equina.
Encéfalo
Rombencéfalo
Mielencéfalo
Es la vesícula encefálica más caudal. Origina el bulbo raquídeo, que difiere de la medula
porque posee movimientos de eversión.
Su placa basal contiene los núcleos motores que se dividen en 3 grupos:
• grupo medial o eferente somático (continuación cefálica de las células del asta anterior –
columna motora eferente somática-. En el mielencéfalo representa las neuronas el XII
par, en el metencéfalo y mesencéfalo esta representado por el III, IV y VI par)
• grupo intermedio o eferente visceral especial (se extiende hasta el metencéfalo
formando la columna motora eferente visceral. Se distribuyen en los músculos estriados
de los arcos faringeos. En el mielencéfalo esta representado por las neuronas del IX, X y
XI par)
• grupo lateral o eferente visceral general (se distribuyen en los músculos lisos del
corazón, aparato respiratorio tracto intestinal).
La placa alar tiene 3 grupos de núcleos sensitivos:
• Grupo somático o lateral (sensitivo. Recibe los impulsos del oído y la superficie de la
cabeza por medio del VIII y V par).
• Grupo intermedio o aferente visceral especial (recibe los impulsos de los botones
gustativos de la lengua y paladar, bucofaringe y epiglotis)
• Grupo medial o aferente visceral general (recibe información interoceptiva del aparato
gastrointestinal y del corazón).
La placa de techo consiste en una capa única de células ependimarias cubierta por piamadre.
Unidas forman la tela coroidea que se introduce en las cavidades ventriculares formando el plexo
coroideo (elabora el LCR)
Metencéfalo
Se encarga de formar el cerebelo (centro de coordinación de la postura y movimientos)y la
protuberancia (vía para las fibras nerviosas entre la medula espinal y la cortezas cerebral y
cerebelosa).
Cada placa basal contiene 3 grupos:
• Grupo eferente somático medial (origen del VI par)
• Grupo eferente visceral especial (contiene núcleos del V y VII par).
• Grupo eferente visceral general (gandulas submaxilares y sublinguales).
La capa marginal de las placas basales se expande y sirve de puente para las fibras nerviosas que
conectan la corteza cerebral con la medula espinal (protuberancia).
Las placas alares contienen 3 grupos:
• Grupo aferente somático lateral (neuronas del V y VIII par)
• Grupo aferente visceral especial
• Grupo aferente visceral general
Cerebelo
Las placas alares se curvan en sentido medial y forman los labios rómbicos que se acercan en la
línea media por debajo del mesencéfalo y forman la placa cerebelosa.
En el embrión de 12 semanas la placa cerebelosa presenta en la línea media el vermis y
lateralmente los hemisferios. Poco después una cisura separa el nódulo del vermis y el floculo lateral
de los hemisferios. Este lóbulo floculonodular es la parte más primitiva del cerebelo.
En una etapa inicial, la placa cerebelosa esta formada por las capas neuroepitelial, del mato y
marginal. Durante el desarrollo ulterior, algunas células formadas por neuroepitelio emigran hacia la
superficie del cerebelo y forman la capa granulosa externa.
En el 6º mes, la capa granulosa externa da origen a las células granulosas, células en cesta y
células estrelladas que comienzan a migrar hacia las células de Purkinje. La corteza del cerebelo esta
formada por células de Purkinje, neuronas Golgi tipo II y neuronas producidas por la granulosa
externas y alcanza sus dimensiones definitivas después del nacimiento. Los núcleos cerebelosos
profundos llegan a su situación definitiva antes del nacimiento.
Mesencéfalo
Es la más primitiva de las vesículas encefálicas (desde el punto de vista morfológico).
Cada placa basal posee 2 grupos:
• Grupo medial o eferente somático (III y IV par)
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
• Grupo eferente visceral general (núcleo de Edinger-Westphal)
La capa marginal de cada placa basal aumenta de tamaño y forma el pie de los pedúnculos
cerebrales.
Las placas alares aparecen en forma de 2 elevaciones longitudinales separadas por una depresión.
Al continuar el desarrollo aparece un surco transversal que divide a cada elevación en un colículo
anterior (superior – tubérculo cuadrigémino superior. Reflejos visuales) y otro posterior (inferior -
tubérculo cuadrigémino inferior. Reflejos auditivos) formados por neuroblastos.
Diencéfalo
PLACA DE TECHO Y EPIFISIS
Se desarrolla a partir de la poción mediana del prosencéfalo. Consiste en una placa de techo y
dos placas alares. Carece de placas de piso y basales.
La placa de techo esta formado por una capa de células ependimarias cubiertas por mesenquima
vascularizado que forma el plexo coroideo del tercer ventrículo. La porción mas caudal de la placa de
techo se convierte en el cuerpo pineal o epífisis (comienza a evaginarse hacia la 7ª semana y se
convierte en un órgano macizo situado en el techo del mesencéfalo).
PLACA ALAR, TALAMO E HIPOTALAMO
Las placas alares forman las paredes laterales del diencéfalo. El surco hipotalamico divide a la
placa en una porción dorsal – tálamo – y una ventral – hipotálamo-. El tálamo sobresale en la luz del
diencéfalo y forma en la línea media la masa intermedia o comisura gris intertalamica.
El hipotálamo forma la porción inferior de la placa alar, se diferencia a varios núcleos que
actúan como centros de regulación del sueño, digestión, temperatura corporal.
HIPOFISIS O GLANDULA PITUITARIA
Se desarrolla en 2 partes:
Una evaginacion ectodérmica del estomodeo, delante de la membrana bucofaríngea (bolsa
de Rathke).
Una prolongación del diencéfalo hacia abajo, el infundíbulo.
Cuando el embrión tiene 3 semanas la bolsa de Rathke se observa como una evaginacion de la
cavidad bucal que crece en sentido dorsal hacia el infundíbulo y hacia el 2º mes se halla en íntimo
contacto con este.
Las células de la pared anterior de la bolsa de Rathke forman la adenohipofisis, la pars tuberalis
crece a lo largo del tallo del infundíbulo y lo rodea. La pared posterior forma la pars intermedia. El
infundíbulo da origen a la pars nervosa o neurohipofisis.
Telencéfalo
Es la vesícula encefálica más rostral, consiste en dos evaginaciones laterales – hemisferios
cerebrales – y una porción mediana – lamina terminal -.
HEMOSFERIOS CEREBRALES
Comienzan a desarrollarse a partir de la 5º semana en forma de evaginaciones bilaterales del
prosencéfalo. Hacia la mitad del 2º mes la porción basal de los hemisferios aumenta de tamaño
sobresaliendo en el interior de los ventrículos laterales y en piso del agujero de Monro, posee un
aspecto estriado y se denomina cuerpo estriado.
En la región donde la pared del hemisferio esta unida al techo del diencéfalo no se producen
neuroblastos, posee una sola capa de células ependimarias cubiertas por mesenquima vascularizado
que en conjunto forman el plexo coroideo, que sobresale en el ventrículo lateral siguiendo una línea
llamada fisura coroidea.
Inmediatamente por encima de la fisura coroidea, la pared del hemisferio esta engrosada
formando el hipocampo, que sobresale en el ventrículo lateral y tiene función olfatoria.
Los hemisferios se expanden y cubren la cara lateral del diencéfalo, el mesencéfalo y la porción
cefálica el metencéfalo.
El cuerpo estriado se expande hacia atrás y se divide en dos partes: el núcleo caudado
(dorsomedial) y el núcleo lenticular (ventrolateral). Esta división se produce por los cilindroejes que
van a los hemisferios, formando un fascículo fibroso que se denomina capsula interna. La pared medial
del hemisferio y la pared lateral el diencéfalo se fusionan, quedando en íntimo contacto el núcleo
caudado y el tálamo.
El continuo crecimiento de los hemisferios cerebrales en dirección anterior, dorsal e inferior
origina los lóbulos frontal, temporal y occipital. La zona situada entre los lóbulos fontal y temporal se
hunde y forma la ínsula de Reil.
DESARROLLO DE LA CORTEZA
La corteza cerebral se desarrolla a partir del palio, que puede dividirse en 2 regiones:
Paleopalio o arquipalio (lateral al cuerpo estriado)
Neopalio (entre el hipocampo y el paleopalio), donde neuroblastos emigran debajo de la
piamadre para diferenciarse a neuronas maduras y, cuando llegan nuevos neuroblastos, los de
la etapa inicial se sitúan en un plano profundo de la corteza.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
La corteza motora posee abundantes células piramidales y las áreas sensitivas se caracterizan por
células granulosas.
COMISURAS
En el adulto, los hemisferios derecho e izquierdo están conectados por las comisuras que
atraviesan la línea media. La más importante utiliza la lámina terminal. La primera en aparecer es la
comisura anterior, conecta el bulbo olfatorio y el área cerebral correspondiente de un hemisferio con
los del lado opuesto. La segunda en aparecer es la comisura del hipocampo o del trígono, sus fibras
nacen en el hipocampo y convergen en la lámina terminal, cerca del techo del diencéfalo. La comisura
más importante es el cuerpo calloso, aparece hacia la 10º semana, comunica áreas no olfatorias de la
corteza derecha e izquierda y se dispone a manera de arco sobre el techo de diencéfalo.
Además de las tres comisuras que aparecen en la lámina terminal, se desarrollan otras tres: dos de
ellas – comisuras posterior y la habenular – se encuentran por delante y por debajo del tallo de la
epífisis. La tercera, el quiasma óptico, aparece en la pared rostral del diencéfalo.
Nervios craneanos
Hacia la 4º semana de desarrollo aparecen los núcleos de los 12 nervios craneanos. Todos ellos,
excepto el I y II par se originan en el tronco del encéfalo y de estos, únicamente el III par nace
fuera de la región del cerebro posterior. En el cerebro posterior los centros de proliferación se
encuentran en el neuroepitelio y establecen 8 rombómeras (segmentos) que originan los núcleos
motores de los pares IV, V, VI, VII, VIII, IX, X, XI y XII.
Las neuronas motoras para los núcleos craneanos se encuentran dentro del tronco del encéfalo,
mientras que los ganglios sensitivos están situados fuera de este.
El origen de los ganglios sensitivos se encuentra en las placodas ectodérmicas (placodas nasales,
auditivas y 4 placodas epibranquiales – engrosamientos ectodérmicos de los arcos faringeos- que
contribuyen a los ganglios de los pares V, VII, IX y X) y en las células de la cresta neural (originan
los ganglios parasimpáticos y sus fibras son conducidas por los pares III, VII, IX y X)
Sistema nervioso autónomo
Puede dividirse en dos partes: el simpático (situado en la región toracolumbar) y el
parasimpático (situado en la región cefálica y sacra).
Sistema nervioso simpático
En la 5º semana de desarrollo la células originarias de la cresta neural de la región torácica
emigran a cada lado de la medula espinal, por detrás de la aorta dorsal, formando una cadena bilateral
de ganglios simpáticos segmentarios conectados entre si por fibras nerviosas longitudinales. Desde su
posición en el tórax, los neuroblastos emigran hacia las regiones cervical y lumbosacra de modo que la
cadenas simpáticas se extienden en toda la longitud de la columna.
Los neuroblastos que emigran por delante de la aorta forman los ganglios preaorticos (celiacos,
mesentéricos), los que se dirigen hacia el corazón, pulmones y aparato gastrointestinal originan los
plexos viscerales simpáticos.
En el asta intermedia de los segmentos toracolumbares de la medula se originan fibras
nerviosas que penetran en los ganglios de las cadenas. Algunas, establecen sinapsis en el mismo nivel
de las cadenas simpáticas o pasan a través de ellas hasta los ganglios aorticos o laterales y se
denominan fibras preganglionares (poseen vaina de mielina y estimulan a las células de los ganglios
simpáticos. Pasan de los nervios raquídeos a los ganglios simpáticos formando los ramos comunicantes
blancos que se extienden de T1 a L2 o L3)
Los cilindroejes de las células ganglionares simpáticas se denominan fibras posganlionares y
carecen de vaina de mielina. Se dirigen hacia otros niveles de la cadena simpática o hacia el corazón,
pulmones y el aparato gastrointestinal (líneas entrecortadas).
Otras fibras, llamadas ramos comunicantes grises van desde la cadena simpática hacia los vasos
sanguíneos periféricos, pelos y glándulas sudoríparas. Estos ramos se encuentran en todos los niveles
de la medula.
GLANDULA SUPRARRENAL
Se desarrolla a partir de 2 componentes:
Una porción mesodérmica que forma la corteza.
Una porción ectodérmica que origina la medula
Durante la 5º semana, las células mesoteliales situadas entre la raíz del mesenterio y la gónada
en desarrollo comienzan a proliferar y se introducen en el mesenquima subyacente, donde se
diferencian a órganos acidófilos que forman la corteza fetal o primitiva de la glándula suprarrenal. Otro
grupo de células que provienen del mesotelio envuelven a la masa celular acidófila y forman la corteza
definitiva de la glándula. Después del nacimiento, la corteza fetal experimenta una regresión rápida,
excepto en su capa mas externa que se convierte en zona reticular.
Mientras se esta formando la corteza fetal, las células originaras del sistema simpático (células
de la cresta neural) invaden su cara medial donde se disponen en cordones y acúmulos dando origen a
la medula suprarrenal. Estas células son cromafines y el único grupo que persiste en el adulto se
encuentra en la medula.
Sistema nervioso parasimpático
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
Las neuronas situadas en el tronco encefálico y la región sacra de la medula espinal dan origen
a fibras parasimpaticas preganglionares. Las fibras de los núcleos del tronco encefálico discurren por el
III, VII, IX y X par. Las fibras posganglionares se originan en neuronas derivadas de la cresta neural y se
dirigen hacia las estructuras que inervan.
SISTEMA RESPIRATORIO
Formación de los esbozos pulmonares
- El divertículo respiratorio (esbozo pulmonar) aparece a la cuarta semana como una evaginación de
la pared ventral del intestino anterior.
- El epitelio de revestimiento de la laringe, tráquea y bronquios y el de los pulmones tiene origen
endodérmico mientras que los componentes cartilaginoso, muscular y conectivo de la tráquea y
pulmones derivan del mesodermo esplácnico.
- Inicialmente el esbozo pulmonar se comunica con el intestino anterior hasta que el divertículo se
extiende caudalmente y queda separado del intestino anterior por los rebordes traqueoesofágicos,
que se fusionan para formar el tabique traqueoesofágico, dividiendo así al intestino anterior en una
porcion dorsal, esófago, y otra ventral, la tráquea y esbozos pulmonares.
- El primordio respiratorio sigue en contacto con la faringe por medio del orificio laríngeo
Laringe
- Su revestimiento interno es de origen endodérmico, pero los cartílagos y músculos derivan de los
arcos faríngeos cuarto y sexto.
- Proliferan rápidamente los cartílagos y el epitelio obstruyendo temporalmente la luz del orificio
laríngeo.
- Se forman recesos laterales, ventrículos laterales, en donde hay tejido que se diferencia en cuerdas
vocales falsas y verdaderas.
Tráquea, Bronquios y Pulmones
- En la quinta semana los esbozos bronquiales (evaginaciones laterales del esbozo pulmonar) que
forman los bronquios principales derecho e izquierdo.
- Los espacios para los pulmones, canales pericardioperitoneales, que son ocupados gradualmente
por los esbozos pulmonares en crecimiento.
- El mesodermo que recubre la parte externa del pulmón se convierte en pleura visceral, y la que
cubre el interior de la pared del cuerpo forma la pleura parietal.
Maduración de los pulmones
- Hasta el séptimo mes los bronquiolos se dividen continuamente en conductos cada vez mas
pequeños y la vascularización aumenta de manera constante.
- Células de los bronquiolos respiratorios se hacen mas delgadas y planas (células epiteliales
alveolares de tipo I) en relación con capilares sanguíneos y linfáticos y los espacios rodeados por
ellas son los alvéolos primitivos o sacos terminales.
- Séptimo mes ya hay muchos capilares como para que haya un normal intercambio de gases. De
aquí hasta luego del nacimiento aumentan en número la cantidad de sacos terminales.
- Antes del nacimiento no se observan alvéolos maduros.
- Al final del sexto mes aparecen las células epiteliales alveolares de tipo II, que producen el
surfactante.
- Los movimientos respiratorios del feto comienzan antes del nacimiento y ocasionan la aspiración
del líquido amniótico.
- El crecimiento de los pulmones en el período posnatal se debe principalmente al aumento del
número de bronquíolos respiratorios y alvéolos y no al incremento del volumen de los alvéolos.
Durante los 10 primeros años de vida se forman nuevos alvéolos.
Embriología del Sistema Muscular
A) Generalidades
• Se desarrolla a partir de la HOJA MESODÉRMICA (excepto ciertos músculos lisos).
• Músculo Esquelético: del MESODERMO PARAXIAL. Que forma:
1. Somitas del occipital al sacro
2. Somitómeros en la cabeza
• Músculo Liso: a partir de la…
1. Hoja esplácnica del mesodermo (que rodea al intestino)
2. Ectodermo (musc. del iris, gl. mamaria y sudorípara)
• Músculo Cardíaco: del MESODERMO ESPLÁCNICO q’ rodea al tubo cardíaco.
B) Músculo Estriado Esquelético
• Somitas y Somitómeros forman los músculos del esqueleto axial, paredes corporales, extremidades
y cabeza.
• Cada somita se diferencia en Esclerotoma, Dermatoma y 2 regiones formadoras de músculo
(Dorsomedial y Dorsolateral)
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
• La Dorsolateral (q’ posee el gen MYO-D) proporciona las cels progenitoras de los musc. de las
extremidades y cabeza
• La Dorsomedial (q’ posee el gen MYF-S) forma el MIOTOMA q’ origina la musculatura epimérica.
• La diferenciación incluye:
1. Fusión de Mioblastos (p/formar fibras largas multinucleadas)
2. Aparición de miofibrillas en el citosol
3. Aparición de estrías al “3er MES”
• Los somitómeros son 7 (“rostrales” a los somitas occipitales), sufren un proceso análogo pro
conservan su organización LAXA y NUNCA se segregan en esclerotomas y dermomiotomas
C) Regulación Molecular
• Los genes q’ regulan el desarrollo son: BMP-4, FGF (de la lámina lateral), Proteínas WNT (del
mesodermo lateral).
• Éstos hacen que las cels dorsolaterales del somita expresen el gen MYO-D
• El ectodermo suprayacente secreta BMP-4 q’ induce la síntesis de prot. WNT por el tubo neural.
• Esto hace q las cels dorsomediales activen el MYF-S.
• Los genes de la flia MYO-D actúan como factores de trascripción.
D) Patrones Formación del Músculo
• Son controlados x el tej. Conjuntivo del lugar hacia donde migran los mioblastos.
• Estos tejidos conjuntivos derivan:
1. Reg cefálica: de la cresta neural
2. Reg. Cervical y occipital: del mesodermo somítico
3. Pared corporal y extremidades: del mesodermo somático.
E) Derivados de los Precursores de Cels Musculares
• En la “5ta SEMANA” las cels están agrupadas en 2 grupos:
1. Epímero: dorsal, de las cels dorsomediales del somita (reorganizado como miotomas)
2. Hipómero: ventral, de las cels dorsolaterales, más grandes
• Inervación:
1. Epímeros: Ramos primarios dorsales
2. Hipómeros: Ramos primarios ventrales
• Del Epímero derivan: Musc. extensores de la columna.
• Del Hipómero derivan: músculos…
1. Cervicales: escaleno, genihioideo, prevertebrales, infrahioideos
2. Torácicos: intercostales ext, int y profundos
3. Abdominales: Musc. anchos y recto mayor
4. Lumbares: cuadrado lumbar
5. Sacros y coccígeos: diafragma pelviano, estriado del ano
F) Músculos de la Cabeza
• Todos los musc. voluntarios cefálicos derivan del Mesodermo PARAXIAL
• Excepto: m. del iris (q’ derivan del ectodermo de la cúpula óptica)
• Tabla q’ representa los orígenes de los musc. craneofaciales
Somitómer
Músculos Par craneal
o
Recto sup, int e inf del ojo. III
1, 2
Laríngeos intrínsecos X
3 Oblicuo Superior IV
4 Musc mandíbula (cierre) V
5 Recto externo VI
6 Musc. Mandíbula (abrir) y 2do arco VII
7 Estilofaríngeo IX
2, 5 (grupo occip) Musc lengua XII
Obs: p/ recordar la relación entre el somitómero y la inervación se puede sumar uno al nro de somitómero
en la mayoría
G) Músculos de las Extremidades
• En la “7ma SEMANA” aparecen los 1ros indicios de formación
• Aparecen en forma de condensación del mesénquima cerca del esbozo
• El mesénquima deriva de las cels dorsolaterales de los somitas
• A medida q’ se alarga el esbozo, el tej se desdobla en flexor y extensor
• Inervación: Los ramos 1ros ventrales (p/ el hipómero) se introducen en el mesénquima del esbozo.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
• Las ramas dorsales y ventrales penetran aisladas pro luego se unen
• N. Radial (p/extensores): combinación de ramas segmentarias dorsales
• N. Cubital (p/flexores): combinación de ramas ventrales
• Es un requisito previo para la diferenciación funcional completa que haya intimo contacto entre el
nervio y las fibras musc.
• P/ dermatomas: los n. raquídeos suministran inervación.
H) Músculo Cardíaco
• De la hoja esplácnica del mesodermo q’ rodea al tubo cardiaco endotelial
• Mioblastos se unen x uniones especiales (futuros discos intercalares)
• Los mioblastos no se fusionan como el musc esquelético
• Algunos fascículos poseen miofibrillas con distribución irregular q’ constituyen las Fibras de
purkinge del sistema excitoconductor
I) Músculo Liso
• Del Ap. Digestivo ----------› Mesodermo esplácnico q’ rodea dichas estructuras
• Vascular -------------------› Mesodermo adyacente al endotelio vascular
• Esfínter, dilatador de la pupila, gl mamaria y sudorípara ----------› ectodermo
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
Sistema Tegumentario
El origen de la piel es doble:
• La capa superficial, la epidermis, proviene del ectodermo superficial.
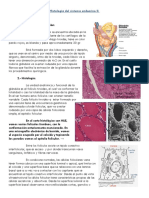
• La capa profunda, la dermis, del mesénquima subyacente.
Epidermis
Al comienzo el embrión tiene una única capa de células ectodérmicas. Al comienzo del 2do. mes, se
divide y forma una capa superficial llamada peridermo o epitriquio (suele desprenderse durante la
segunda mitad de la vida intrauterina). Luego, se forma una tercera capa entre las últimas, la zona o capa
intermedia.
Al final del 4to mes se forman las 4 capas definitivas:
• Estrato basal: capa proliferativa.
• Estrato espinoso: celúlas poliédricas.
• Estrato granuloso: contiene gránulos de queratohialina.
• Estrato córneo: celúlas muertas y compactas.
Durante los 3 primeros meses la epidermis es invadida por células originadas en la cresta neural que
sintetizan melanina (produce la pigmentación de la piel después del nacimiento).
Dermis
Deriva del mesénquima. Durante el tercero y cuarto mes este tejido, llamado corion origina las
papilas dérmicas que sobresalen hacia la epidermis.
La capa más profunda de la dermis, subcorion, contiene abundante tejido adiposo.
En el neonato la piel está cubierta por vernix caseosa o unto sebáceo (secreción de las glándulas
sebáceas y células epidérmicas y pelos degenerados) que la protege contra la maceración por el líquido
amniótico.
Pelo
Aparecen por proliferaciones epidérmicas que se introducen en la dermis. En el extremo terminal los
brotes pilosos se invaginan y forma las papilas pilosas. Luego, las células del centro de los brotes pilosos
se queratinizan y constituyen el tallo del pelo y las células periféricas dan origen a la vaina pilosa epitelial.
La vaina radicular dérmica y el músculo erector del pelo derivan del mesénquima.
Al final del 3er. mes aparecen los primeros pelos en la superficie, en la región de las cejas y el labio
superior. Estos pelos constituyen el lanugo que se desprende antes del nacimiento.
Glándulas mamarias
Al comienzo se produce un engrosamiento a manera de banda en la epidermis, la línea mamaria o
pliegue mamario, que en el embrión de 7 semanas se extiende de la extremidad superior a la inferior.
En este sitio se forman de 16 a 24 brotes que hacia el final de la vida intrauterina se canalizan y
forman los conductos galactóforos. Las evaginaciones constituyen los conductos de menor calibre y los
alvéolos de la glándula.
Patologías
Ictiosis: excesiva queratinización de la piel.
Hipertricosis: exceso de pelo.
Atriquia: falta congénita de pelo.
Politelia: pezones accesorios.
Polimastia: mamas supernumerarias
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
APARATO UROGENITAL
Desde el punto de vista funcional se dividen en:
- Aparato urinario
- Aparato genital
Desde el punto de vista embriológico y anatómico están en íntima relación porque ambos derivan del
mesodermo intermedio.
APARATO URINARIO
Sistemas Renales:
Pronefros: Rudimentario
- Aparece a inicios de la 4ta semana en la región cervical del mesodermo intermedio.
- Formados por 7 a 10 cúmulos celulares. Forman las neuronas (unidades excretoras vestigiales).
- Desaparecen totalmente al final de la 4ta semana.
Mesonefros y conductos mesonéfricos: Funciona poco tiempo.
- Aparecen en la 4ta semana (durante la regresión del sistema paranéfrico) en los segmentos
torácicos y lumbares superiores del mesodermo intermedio.
- Primero: Aparecen los túbulos excretores del mesodermo.
Los túbulos se alargan y reciben a un plexo de capilares (glomérulo).
Forman la cápsula de Bowman
- En el extremo opuesto de este túbulo desemboca en el conducto mesonéfrico de Wolf.
- En la mitad del 2do mes:
- Se forma la Cresta Urogenital - Mesonefro (órgano ovoide voluminoso).
- Gónadas en el lado interno del mesonefro.
- Desaparecen: Primero: Segmento craneal.
Segundo: Segmento caudal. (Persiste en el varón: aparto genital).
Metanefros: Forma el riñón definitivo.
- Aparecen en la 5ta semana.
- Tiene unidades excretoras: derivan del mesodermo metanéfrico.
colectoras: derivan del brote ureteral (evaginación del conducto mesonéfrico
cerca de su desembocadura en la cloaca).
Sistema Colector: (del brote ureteral)
- Forma la pelvis. Este se divide en uno de sus extremos y forma los cálices mayores. Éstos se
vuelven a dividir en 12 generaciones o más.
- La 2ª, 3ª y 4ª generaciones: cáliz menor.
- La 5ª y sucesivas generaciones: túbulos colectores (1 a 3 millones).
- Pirámide renal: Convergencia de varios túbulos colectores en el cáliz menor.
- En el otro extremo de la pelvis se continúa como uréter.
Sistema Excretor: Se forma a partir de células de tejido metanéfrico.
Primero forman vesículas, luego túbulos.
Nefronas
Parte Proximal Parte Distal:
Cápsula de Bowman - Desemboca en
+ glomérulo (capilares) túbulos colectores
- Al nacer hay un millón de nefronas.
- La producción de orina se inicia poco después de la formación de los capilares (10ª semana).
POSICIÓN DEL RIÑÓN
Primero: pélvico
Asciende por disminución de la curvatura del cuerpo.
Segundo: lumbar crecimiento de la región lumbar y sacra.
- En la región pélvica: irrigado por una rama pélvica de la aorta.
- En la región lumbar: irrigado por ramas de la aorta que nacen a niveles más altos.
FUNCIÓN DEL RIÑÓN
- Funciona al final del primer trimestre (aproximadamente en la 12ª semana).
- Pero lo que orina no es la excreción de productos de deshechos (función de la placenya), sino una
deglución del líquido amniótico. Es decir; el feto traga y orina líquido amniótico.
VEJIGA Y URETRA
Desde la 4ta a la 7ma semana: la cloaca se divide en: - seno urogenital (adelante).
- Conducto urorrectal (atrás)
Entre ambos:
Una capa de mesodermo: tabique urorrectal
Este tabique formará el cuerpo perineal.
Seno Urogenital: 3 porciones:
1. Vejiga urinaria: Primero se continúa con la alantoides, después esta se oblitera y la vejiga se une
al ombligo a través del uraco (ligamento umbilical medio).
2. Porción pélvica del seno urogenital: En el varón origina la uretra prostática y membranosa.
3. Porción fálica: Muy aplanada y se desplaza ventralmente porque crece el tubérculo genital. Su
desarrollo difiere según el sexo.
Los conductos mesonéfricos y el brote ureteral desembocan en la vejiga y se separan.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
Desciende más y forma en el varón: Conductos eyaculadores.
La mucosa de la vejiga en la porción en donde desembocan los conductos (trígono vesical) es primero de
origen mesodérmico, pero luego es reemplazado por epitelio endodérmico.
URETRA
Epitelio de la uretra: Endodermo.
Tejido conectivo y muscular: Hoja esplácnica del mesodermo.
Final del 3er. mes: El epitelio prolifera y forma evaginaciones que se introducen en el mesémquima.
Estos brotes originan En el varón: glándula prostática.
En la mujer: glándulas uretrales y parauretrales.
APARATO GENNITAL
La clave del diformismo sexual es el cromosoma Y que tiene el gen SRY (región determinante del sexo
en el cromosoma Y) en el su brazo corto (Yp11). El producto proteíco del gen: factor determinante
testicular.
- Factor Determinante testicular positivo: Sentido Masculino.
- Factor Determinante Testicular negativo: Sentido Femenino.
GÓNADAS
- Determinación genética del sexo: en el momento de la fecundación.
- Gónadas con carácterísticas màsculino o femenino: 7ma semana.
- Primero: Gónadas son solo eminencias longitudinales: pliegues o crestas genitales o gonadas. Luego: las
células germinales aparecen en los pliegues genitales.
- Las célulars germinales primordiales primero aparacen en la pared del saco vitelino cerca de la
alantoides, luego emigran hasta llegar a las gónadas primitivas (5ta. semana). Y ahí invaden a las crestas
genitales (6ta. semana).
- Antes de la llegada de las células germinales primordiales, el epitelio de la cresta genital prolifera
y penetra en el mesénquima subyacente, formando así los cordones sexuales primitivos.
- Pero como el epitelio sigue unido a l superficie, no se diferencia si la gónada es masculina o
femenina. Entonces se la llama gónada indiferente.
TESTÍCULO
Si hay factor determinante testicular (gen SRY) los cordones sexuales siguen proliferando y forman los
cordones testiculares o medulares.
Estos cordones se disgregan en una red de diminutos filamentos: red de Haller o rete testis.
Luego se forma la túnica albugínea (tej conectivo fibroso) que separa los cordones testiculares del
epitelio superficial
En el 4to mes: Cordones testiculares formados por células germinales primordiales y células de Sertoli
(derivada del epitelio superficial de la glándula).
Las células de Leydig:
- Se desarrolla del mesénquima de la cresta genital poco después de la diferenciación de los cordones.
- Están entre los cordones testiculares.
- Producen testosterona (8va semana) y ésta induce a la diferenciación de los conductos genitales y de
los genitales externos.
- En la pubertad: Cordones testiculares originan a los túbulos seminíferos. Estos se continuan con la
red de Haller y estos a su vez con los conductillos eferentes. Éstos son restos de los túbulos
mesonéfricos que unen a la red de Haller y al conducto mesonéfrico llamado Conducto Deferente.
OVARIO
- Como no hay factor determinante testicular, los cordones primitivos se disgregan en cúmulos
celulares que contienen células germinales y se situan en la médula del ovario. Estos cúmulos son
sustituidos por estroma vascularizada, la médula ovárica.
- 7ma semana: El epitelio superficial de la gónada de origen a los cordones corticales (2da
generación de cordones) que penetra en el mesénquima.
- En el 4to mes:
Ovogonias
Células foliculares
Los cordones corticales se disgregan en cúmulos celulares, cada uno alrededor de una o más células
germinales primitivas.
El sexo genético del embrión es determinado genéticamente en la fecundación y depende de que el
espermatozoide tenga un cromosoma X o Y.
Embriones XX: Los cordones medulares desaparecen y se desarrollan los cordones corticales.
Embriones XY: Los cordones medulares se convierten en cordones testiculares y no se desarrolan los
cordones corticales.
CONDUCTOS GENITALES
Período indiferenciado:
Embriones masculinos y femeninos tienen al inicio dos pares de conductos genitales: los conductos
mesonéfricos (o de Wolf) y los conductos paramesonéfricos (o de Müller).
• El conducto paramesonéfrico (invaginación de la cresta longitudinal):
- En sentido craneal: Desemboca en la cavidad abdominal.
- En sentido caudal: 1ro: Situado externo al conducto mesonéfrico
2do: Cruza al conducto mesonéfrico ventralmente.
3ro: Se une en la línea media con el conducto del lado opuesto.
4to: Se fusionan y forman el conducto uterino.
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
5to: Desemboca en la pared posterior del seno urogenital, donde forma un
abultamiento, el tubérculo paramesonéfrico o de Müller.
• Los conductos mesonéfricos desembocan en el seno urogenital, a cada lado del tubérculo de Müller.
Conductos genitales masculinos: sdfadfas
• Túbulos epigenitales (túbulos excretores mesonéfricos que no sufrieron regresión) se unen con
los cordones de la red de Haller y forman los conductillos eferentes del testículo.
• Los túbulos excretores del polo caudal del testículo no se unen a la red de Haller, son los túbulo
paragenitales. Los vestigios de estos conductillos paradídimo.
Conductos mesonéfricos En su porción craneal: Desaparecen y forman el
apéndice del epidídimo o idátide pediculada.
En su porción caudal: Persisten. Se alargan y se arrollan entre sí,
formando el epidídimo.
• Desde el epidídimo hasta la vesícula seminal: Conductos mesonéfricos adquieren una gruesa túnica
muscular y se convierten en el conducto deferente.
• Después de la vesícula seminal se denomina conducto eyaculador.
• En el varón, el conducto paramesonéfrico degenera excepto en la porción craneal que forma el
apéndice del testículo o hidátide sésil (de Morgagni).
Conductos genitales femeninos:
• Derivan de los conductos paramesonéfricos.
• Tienen 3 porciones: a) Craneal vertical: Desemboca en la cevidad abdominal.
b) Horizontal: Cruza ventralmente el conducto mesonéfrico.
c) Caudal Vertical: Se fusiona con el del lado opuesto.
• Al descender el ovario, las dos primeras forman la trompa uterina o de falopio y las partes
caudales fusionadas forman el conducto uterino que dará origen al cuello y al cuerpo del
útero. (Éstos rodeados por una capa de mesénquima, que es la capa muscular del útero o
miometrio y su revestimiento peritoneal, el perimetrio).
• Después de la fusión de los conductos en la línea media, se crea un repliegue pelviano transversal
ancho que va desde la parte externa de los conductos fusionados hasta la pared pelviana. Es el
ligamento ancho del útero.
• El útero y los lig. anchos dividen a la pelvis en: - Fondo de saco uterorrectal
- Fondo de saco vesicouterino
VAGINA
• Doble origen Superior: Conducto uterino.
Inferior: Seno urogenital.
• Del seno urogenital se extienden evaginaciones bulbos sinovaginales
Forma una lámina vaginal
(porción inferior y media de la vagina)
• Las prolongaciones que se disponen a manera de alas de la vagina alrededor del extremo del útero
son los fondos de saco vaginales. Éstos forman junto con el útero la cúpula vaginal (porción
sueprior de la vagina), que es de origen promesonéfrico.
• Himen: Separa el interior del seno urogenital del interior de la vagina. Formado por el
revestimiento epitelial del seno y una delgada capa de células vaginales.
• Epoóforo y paraoóforo: Restos de túbulos excretores craneales y caudales, respectivamente, en
el mesoovario.
• Generalmente estos desaparecen, excepto una porción craneal (en el epoóforo) y a veces, una
porción caudal (en el útero o en la vagina).
Este podría formar más adelante el quiste de Gartner.
GENITALES EXTERNOS
Período indiferenciado:
3ra semana: Células mesenquimáticas migran y forman eminencias alrededor de la membrana cloacal, los
pliegues cloacales.
Estos pliegues___Craneal a la memb cloacal: Se unen y forman tubérculo genital
Caudal a la memb cloacal: Se subdividen ___Pliegues uretrales
Pliegues anales
A cada lado de los pliegues uretrales: eminencias genitales.
En el varón: eminencias escrotales.
En la mujer: labios mayores.
Genitales externos masculinos: Bajo la influencia de los andrógenos
• Alargamiento del tubérculo genital que ahora se llama falo.
• Al alargarse, el falo tira hacia delante los pliegues uretrales y estos forman las paredes laterales del
surco uretral, que se extiende a lo largo de la cara caudal del falo, pero no llega hasta el glande.
• Este surco está revestido por la lámina uretral (epitelio de origen endodérmico)
• Final del 3er mes: Pliegues uretrales se cierran sobre la lámina uretral y forman la uretra peniana.
Pero como el surco no llega has ta el glande, la porción distal de la uretra se forma cuando las
células ectodérmicas de la punta del glande se dirigen hacia adentro y forman un cordón corto (4to
mes).
Descargado por Lisette Juares (juareslisette5@gmail.com)
lOMoARcPSD|4000730
• Este corto cordón se canaliza y forma el meato uretral externo.
• Las eminencias escrotales:
1ro: En la región inguinal
2do: Se desplazan caudalmente y cada una forma la mitad del escroto (separadas por el tabique o
rafe escrotal).
Genitales externos femeninos: Estimulados por estrógenos.
• Tubérculo genital se alarge apenas un poco y forma el clítoris. (Pero el tubérculo genital es más
grande que en el varón en los primeros períodos del desarrollo).
• Los pliegues uretrales no se fusionan sino que se transforman en los labios menores.
• Las eminencias genitales se agrandan y forman los labios mayores.
• El surco urogenital queda abierto y forma el vestíbulo.
Descargado por Lisette Juares (juareslisette5@gmail.com)
Potrebbero piacerti anche
- Resumen LangmanDocumento20 pagineResumen LangmanGabrielaRojas60% (5)
- Resumen LangmanDocumento29 pagineResumen Langmanemergencias275200491% (11)
- Resumen Ciclo Ovarico PDFDocumento5 pagineResumen Ciclo Ovarico PDFJOSÉ MIGUEL ANTONIO SOTO ROJAS0% (1)
- Embriologia Ojo-Oido Clase IDocumento46 pagineEmbriologia Ojo-Oido Clase IAnonymous UYKHA42Nessuna valutazione finora
- Preguntas Primera Semana EmbriologiaDocumento1 paginaPreguntas Primera Semana Embriologiahumberto100% (2)
- Resumen de EmbriologiaDocumento40 pagineResumen de Embriologiajonathan288986% (7)
- Examen EmbrioDocumento13 pagineExamen EmbrioMedicina UiC100% (5)
- 06 Embriologia FinalDocumento26 pagine06 Embriologia FinalkaritoallrigtNessuna valutazione finora
- Resumen Capitulo 12 Apuntes Muy Completos Del Langman PDFDocumento6 pagineResumen Capitulo 12 Apuntes Muy Completos Del Langman PDFtania ponceNessuna valutazione finora
- Resumen de RossDocumento188 pagineResumen de RossRaul Melendez Navarro100% (13)
- Fisiología de Guyton Resumen Capítulo 8Documento2 pagineFisiología de Guyton Resumen Capítulo 8Pablo Andrés Zurita86% (21)
- Cuestionario EmbriologiaDocumento2 pagineCuestionario EmbriologiaangelicaNessuna valutazione finora
- Sistema Humoral Del OjoDocumento4 pagineSistema Humoral Del Ojoapi-457710535Nessuna valutazione finora
- Embriología Del Sistema CardiovascularDocumento3 pagineEmbriología Del Sistema Cardiovascularolivia2390100% (1)
- Histologia Del Sistema EndocrinoDocumento12 pagineHistologia Del Sistema EndocrinoKarla Gabriela Villegas JibajaNessuna valutazione finora
- Desarrllo Aparato Urogenital San MartinDocumento42 pagineDesarrllo Aparato Urogenital San MartinUSMP FN ARCHIVOSNessuna valutazione finora
- Resumen Histologia RossDocumento9 pagineResumen Histologia RosshongdanNessuna valutazione finora
- Apuntes de EmbriologiaDocumento29 pagineApuntes de EmbriologiaDiego Armando Arancibia Bernal67% (3)
- Sistema Urogenital (Guía de Actividad Práctica)Documento2 pagineSistema Urogenital (Guía de Actividad Práctica)Heylling SoteloNessuna valutazione finora
- Fisiologia D Guyton ResumenDocumento17 pagineFisiologia D Guyton ResumenJesús Alberto TatisNessuna valutazione finora
- Embriologia de Sistema CardiovascularDocumento65 pagineEmbriologia de Sistema Cardiovascularapi-2681888577% (13)
- Histologia Del Sistema Endocrino IIDocumento12 pagineHistologia Del Sistema Endocrino IIPedro GonzalezNessuna valutazione finora
- AnencefaliaDocumento3 pagineAnencefaliaKasamivega100% (1)
- Histologia Del Sistema Reproductor FemeninoDocumento53 pagineHistologia Del Sistema Reproductor FemeninoMiLita Cruz Trujillo100% (1)
- Histologia CuestionarioDocumento28 pagineHistologia CuestionariochevelosoNessuna valutazione finora
- Embriologia Usmp SeminarioDocumento9 pagineEmbriologia Usmp SeminarioLuz Valderrama DurandNessuna valutazione finora
- VejigaDocumento26 pagineVejigaVane San50% (2)
- Cabeza Resumen Del Libro de Moore de Anatomia Humana PDFDocumento34 pagineCabeza Resumen Del Libro de Moore de Anatomia Humana PDFRevisa RevisaNessuna valutazione finora
- S Urinario Ross 2009Documento9 pagineS Urinario Ross 2009JA Torres100% (5)
- Procesos de GestaciónDocumento67 pagineProcesos de Gestaciónleondario223hotmailcNessuna valutazione finora
- Desarrollo de Cavidades CorporalesDocumento2 pagineDesarrollo de Cavidades CorporalesArantza CuenNessuna valutazione finora
- Tercera Semana Embriologia HumanaDocumento33 pagineTercera Semana Embriologia HumanaJhonatan Israel Freire Liquín100% (3)
- Histologia Aparato Genital MasculinoDocumento5 pagineHistologia Aparato Genital Masculinocheveloso100% (2)
- Fisiología de Guyton Resumen Capítulo 1Documento2 pagineFisiología de Guyton Resumen Capítulo 1Pablo Andrés Zurita92% (25)
- Histología Aparato UrinarioDocumento51 pagineHistología Aparato UrinarioAlex Yvan Escobedo Hinostroza100% (4)
- Capitulo 18 Resumen de Sistema Nervioso Central Langman 12Documento14 pagineCapitulo 18 Resumen de Sistema Nervioso Central Langman 12raul50% (4)
- Histología II Examen FinalDocumento85 pagineHistología II Examen FinalJohnRudyHuallpaVila0% (1)
- Preguntas EmbriologíaDocumento8 paginePreguntas EmbriologíaSantiago Anchi0% (1)
- Cavidades CorporalesDocumento9 pagineCavidades CorporalesBelen Yapu100% (1)
- A81 Embriologia Del Aparato DigestivoDocumento79 pagineA81 Embriologia Del Aparato DigestivoTrinidad92% (39)
- Resumen Embriologia LangmanDocumento13 pagineResumen Embriologia Langmanmanuel_aguirre_13Nessuna valutazione finora
- Fisiología de GametogénesisDocumento5 pagineFisiología de Gametogénesispier manuel cabezas armellonNessuna valutazione finora
- Loe11 FisiologiaDocumento30 pagineLoe11 FisiologiaJuan Pablo Giraldo MontoyaNessuna valutazione finora
- Lección No 1. Fisiología de La Reproduccion1Documento29 pagineLección No 1. Fisiología de La Reproduccion1vimarjuNessuna valutazione finora
- GametogénesisDocumento3 pagineGametogénesisivanna camposNessuna valutazione finora
- El Pibe Resumen de FecundaciónDocumento29 pagineEl Pibe Resumen de FecundaciónAntonella Violo100% (1)
- Temas de Embriologia ENURMDocumento35 pagineTemas de Embriologia ENURMLissetteAlexandraPerezNessuna valutazione finora
- Guia Gametogenesis, Fecundacion y 1ra SemanaDocumento5 pagineGuia Gametogenesis, Fecundacion y 1ra SemanaeldoctopuntocomNessuna valutazione finora
- Gametogénesis y Embriología Del Aparato Genital FemeninoDocumento12 pagineGametogénesis y Embriología Del Aparato Genital FemeninoAlma Linda Peréz Santeliz100% (1)
- Embrioapuntes (Parte I) - Diego MeléndezDocumento16 pagineEmbrioapuntes (Parte I) - Diego Meléndezdiego melendezNessuna valutazione finora
- GametogénesisDocumento4 pagineGametogénesisAlexander EztradaNessuna valutazione finora
- EmbriologíaDocumento17 pagineEmbriologíaCarmen Porras Jarufe50% (2)
- Guia Embriologia CeabioDocumento5 pagineGuia Embriologia CeabioHaykareth Mi Amor BelloNessuna valutazione finora
- GametogénesisDocumento25 pagineGametogénesisEsmeraldaNessuna valutazione finora
- Gametogenesis M1.3Documento10 pagineGametogenesis M1.3Hector Efrain Garcia ChavezNessuna valutazione finora
- Resumen de EmbriologíaDocumento26 pagineResumen de EmbriologíaEvelyn Gonzalez100% (1)
- 2 - Origen y Desarrollo Del Embarazo (Capitulo 2)Documento8 pagine2 - Origen y Desarrollo Del Embarazo (Capitulo 2)Roxana Kong75% (4)
- Guía Embriología 1er ParcialDocumento35 pagineGuía Embriología 1er ParcialAlberto FongNessuna valutazione finora
- Reporte de Laboratorio de GametogénesisDocumento14 pagineReporte de Laboratorio de GametogénesisYessy Ballesteros100% (1)
- 21) Miología de Cabeza y CuelloDocumento83 pagine21) Miología de Cabeza y CuelloL JNessuna valutazione finora
- 8) Miembro Superior 2Documento47 pagine8) Miembro Superior 2L JNessuna valutazione finora
- 24) SN Parte I GENERALIDADESDocumento33 pagine24) SN Parte I GENERALIDADESL JNessuna valutazione finora
- Trayecto InguinalDocumento1 paginaTrayecto InguinalL JNessuna valutazione finora
- Obstetricia Normal - Bolilla 20 - Presentación Cefálica de VérticeDocumento4 pagineObstetricia Normal - Bolilla 20 - Presentación Cefálica de VérticeL JNessuna valutazione finora
- Bolilla 19 ALUMBRAMIENTO PDFDocumento4 pagineBolilla 19 ALUMBRAMIENTO PDFL JNessuna valutazione finora
- Obstetricia Normal - Bolilla 18 - Evaluación de La Salud FetalDocumento3 pagineObstetricia Normal - Bolilla 18 - Evaluación de La Salud FetalL JNessuna valutazione finora
- Preguntas de EmbriologiaDocumento36 paginePreguntas de EmbriologiaL JNessuna valutazione finora
- Bolilla 21 PRESENTACIONES DEFLEXIONADAS PDFDocumento5 pagineBolilla 21 PRESENTACIONES DEFLEXIONADAS PDFL JNessuna valutazione finora
- Segunda Semana EmbrioDocumento4 pagineSegunda Semana EmbrioSusan Mel Corani QuelcaNessuna valutazione finora
- Estudiar EmbriologiaDocumento14 pagineEstudiar EmbriologiaEVA YAMILETH FONSECA REYES100% (1)
- EmbriologiaDocumento22 pagineEmbriologiaRebeca RamírezNessuna valutazione finora
- Fecundación, Segmentación e ImplantaciónDocumento23 pagineFecundación, Segmentación e ImplantaciónKimberlyChoqueGallegosNessuna valutazione finora
- Generalidades de Crecimiento y Desarrollo Grupo 2Documento77 pagineGeneralidades de Crecimiento y Desarrollo Grupo 2Daniel Vega AdauyNessuna valutazione finora
- Desarrollo Embriológico Del PorcinoDocumento5 pagineDesarrollo Embriológico Del PorcinoJhojan Victorio ChachiNessuna valutazione finora
- Etapas de La Reproducción para SextoDocumento12 pagineEtapas de La Reproducción para SextoMAILLINVALERONessuna valutazione finora
- EMBRIOLOGIA 1 Primera Semana de Desarrollo de La Ovulacion Ala ImplantacionDocumento18 pagineEMBRIOLOGIA 1 Primera Semana de Desarrollo de La Ovulacion Ala ImplantacionAlejandra celeste Mollo GonzalesNessuna valutazione finora
- Implantacion Embrionaria 7 SDocumento43 pagineImplantacion Embrionaria 7 SALEXANDRANessuna valutazione finora
- Endometritis CrónicaDocumento3 pagineEndometritis CrónicaElizabeth Mendoza EspinosaNessuna valutazione finora
- Embriologia (Carlos Lehmann)Documento74 pagineEmbriologia (Carlos Lehmann)AdriánNessuna valutazione finora
- Embriologia SexualidadDocumento163 pagineEmbriologia SexualidaddavidNessuna valutazione finora
- ImplantaciónDocumento2 pagineImplantaciónRich MtzzNessuna valutazione finora
- Unidad 2 Reproduccion y Sexualidad Guia N 2 3er CicloDocumento16 pagineUnidad 2 Reproduccion y Sexualidad Guia N 2 3er CiclocamiloNessuna valutazione finora
- Desarrollo Del EmbarazoDocumento12 pagineDesarrollo Del EmbarazoalvaroNessuna valutazione finora
- 5 Embriologia PlacentaDocumento55 pagine5 Embriologia PlacentaMarisol GuanoNessuna valutazione finora
- Guia Desarrollo EmbrionarioDocumento7 pagineGuia Desarrollo EmbrionarioFredman Cruz YatNessuna valutazione finora
- Endometriosis MonografiaDocumento36 pagineEndometriosis MonografiaKassandraAltamiranoNessuna valutazione finora
- Desarrollo Embrionario, Fetal y EmbarazoDocumento77 pagineDesarrollo Embrionario, Fetal y EmbarazoMari Gzlez100% (1)
- PRACTICA 08 y 09 PDFDocumento9 paginePRACTICA 08 y 09 PDFNar JovNessuna valutazione finora
- Desarrollo Embrionario FetalDocumento14 pagineDesarrollo Embrionario FetalAnorudo ConamorNessuna valutazione finora
- Transporte e Implantacion Del EmbrionDocumento2 pagineTransporte e Implantacion Del EmbrionClaudia Arellano UrquizoNessuna valutazione finora
- Criopreservación de EmbrionesDocumento46 pagineCriopreservación de EmbrionespercyNessuna valutazione finora
- GUIA SEMANA 7 Part 2Documento19 pagineGUIA SEMANA 7 Part 2Gabriela ColinaNessuna valutazione finora
- Sílabo Embriología 2019-Ii PDFDocumento13 pagineSílabo Embriología 2019-Ii PDFVanessa AvilaNessuna valutazione finora
- Se Expone El Caso de Una Perra Bulldog Inglés Que Presentaba Una Gestación Aproximada de 36 A 38 DíasDocumento9 pagineSe Expone El Caso de Una Perra Bulldog Inglés Que Presentaba Una Gestación Aproximada de 36 A 38 DíasPablo TorizNessuna valutazione finora
- Gastrulacic3b3n Delimitacic3b3n y Hojas Embrionarias UtisDocumento54 pagineGastrulacic3b3n Delimitacic3b3n y Hojas Embrionarias UtisLimber Paño YujraNessuna valutazione finora
- La PlacentaciónDocumento12 pagineLa PlacentaciónKira Tomoki100% (1)
- Biología de La Concepcion, Embriología y Anatomía de Aparato Reproductor FemeninoDocumento26 pagineBiología de La Concepcion, Embriología y Anatomía de Aparato Reproductor FemeninoJoyce de VeraNessuna valutazione finora
- Laboratorio y Embarazo PDFDocumento50 pagineLaboratorio y Embarazo PDFLeslyBubaNessuna valutazione finora