Potrebbero piacerti anche
- Of Maybugs and Men: A History and Philosophy of the Sciences of HomosexualityDa EverandOf Maybugs and Men: A History and Philosophy of the Sciences of HomosexualityNessuna valutazione finora
- Sesardic, N. 2010. "Race: A Social Destruction of A Biological Concept"Documento20 pagineSesardic, N. 2010. "Race: A Social Destruction of A Biological Concept"YmeNessuna valutazione finora
- Human No More: Digital Subjectivities, Unhuman Subjects, and the End of AnthropologyDa EverandHuman No More: Digital Subjectivities, Unhuman Subjects, and the End of AnthropologyValutazione: 1 su 5 stelle1/5 (1)
- Matters of RaceDocumento10 pagineMatters of RaceJoséRamónOrrantiaCavazosNessuna valutazione finora
- Ingenious Nonsense: Religion, Philosophy, Pseudoscience, and the ParanormalDa EverandIngenious Nonsense: Religion, Philosophy, Pseudoscience, and the ParanormalNessuna valutazione finora
- Neither Man Nor Woman - Berdache - A Case For Non-Dichotomous Gender Construction.Documento18 pagineNeither Man Nor Woman - Berdache - A Case For Non-Dichotomous Gender Construction.shubhamrai266Nessuna valutazione finora
- Perspectives on Science and CultureDa EverandPerspectives on Science and CultureKris RuttenNessuna valutazione finora
- SSG1976Documento5 pagineSSG1976Rafaella GaleanoNessuna valutazione finora
- These "Thin Partitions": Bridging the Growing Divide between Cultural Anthropology and ArchaeologyDa EverandThese "Thin Partitions": Bridging the Growing Divide between Cultural Anthropology and ArchaeologyNessuna valutazione finora
- Realism, Antirealism, and Conventionalism About RaceDocumento33 pagineRealism, Antirealism, and Conventionalism About RaceJuan Jacobo Ibarra SantacruzNessuna valutazione finora
- Ethical Universe: the Vectors of Evil Vs. Good: Secular Ethics for the 21St CenturyDa EverandEthical Universe: the Vectors of Evil Vs. Good: Secular Ethics for the 21St CenturyNessuna valutazione finora
- This Content Downloaded From 75.69.132.126 On Thu, 28 Jan 2021 19:10:40 UTCDocumento26 pagineThis Content Downloaded From 75.69.132.126 On Thu, 28 Jan 2021 19:10:40 UTCOrsaNessuna valutazione finora
- Beyond Fact or Fiction On The Materiality of RaceDocumento24 pagineBeyond Fact or Fiction On The Materiality of RaceArturo SanabriaNessuna valutazione finora
- 2002 12 TrendscogsciDocumento2 pagine2002 12 TrendscogsciAdrian GuzmanNessuna valutazione finora
- Sesardic, N. 2013. "Confusions About Race: A New Installment"Documento7 pagineSesardic, N. 2013. "Confusions About Race: A New Installment"YmeNessuna valutazione finora
- Annual Reviews: Info/about/policies/terms - JSPDocumento24 pagineAnnual Reviews: Info/about/policies/terms - JSPPedro LolliNessuna valutazione finora
- Cultural Relativism 2.0Documento22 pagineCultural Relativism 2.0Juno ZhangNessuna valutazione finora
- The University of Chicago Press, Wenner-Gren Foundation For Anthropological Research Current AnthropologyDocumento22 pagineThe University of Chicago Press, Wenner-Gren Foundation For Anthropological Research Current AnthropologySecret SantaNessuna valutazione finora
- 9403093119Documento24 pagine9403093119Huda GüzelNessuna valutazione finora
- Genetics and The Origin of Human "Races"Documento15 pagineGenetics and The Origin of Human "Races"esitgen100% (1)
- No Way Out of The Binary: A Critical History of The Scientific Production of SexDocumento27 pagineNo Way Out of The Binary: A Critical History of The Scientific Production of SexCeciliaRustoNessuna valutazione finora
- MigraciónDocumento82 pagineMigraciónnoehernandezcortezNessuna valutazione finora
- Review of A Troublesome Inheritance by Nicholas WadeDocumento8 pagineReview of A Troublesome Inheritance by Nicholas Wadedaniel leite JrNessuna valutazione finora
- Porqué en Contra de DarwinDocumento10 paginePorqué en Contra de DarwinAlberto Jesús CamposNessuna valutazione finora
- Dawkins and Biological DeterminismDocumento4 pagineDawkins and Biological DeterminismdennistrilloNessuna valutazione finora
- Pertemuan 10 - Claude Lévi-Strauss - Structural Anthropology, Volume 2-Basic Books (1976)Documento415 paginePertemuan 10 - Claude Lévi-Strauss - Structural Anthropology, Volume 2-Basic Books (1976)jengkolmaniaaNessuna valutazione finora
- Selcer 2012Documento13 pagineSelcer 2012Rafaella GaleanoNessuna valutazione finora
- Why Do We Think RaciallyDocumento4 pagineWhy Do We Think RaciallyParag Bhatnagar100% (1)
- Interrogating WhitenessDocumento19 pagineInterrogating WhitenessPerla ValeroNessuna valutazione finora
- Toward A Consilient Study of Literature: Pinker, Steven, 1954Documento18 pagineToward A Consilient Study of Literature: Pinker, Steven, 1954UgartecheNessuna valutazione finora
- 1 s2.0 S2352552522000925 MainDocumento2 pagine1 s2.0 S2352552522000925 MainCharlierNessuna valutazione finora
- Can There Be A Nietzschean SociologyDocumento20 pagineCan There Be A Nietzschean SociologyQisen FuNessuna valutazione finora
- 20101004225016242Documento312 pagine20101004225016242André Cidade100% (2)
- Rossi, Ino - The Unconscious in The Anthropology of Claude Lévi-StraussDocumento30 pagineRossi, Ino - The Unconscious in The Anthropology of Claude Lévi-StraussRoberto Nación PantigosoNessuna valutazione finora
- Is Science RacistDocumento5 pagineIs Science Racistgumel99Nessuna valutazione finora
- PodcastDocumento9 paginePodcastKevin YCDNessuna valutazione finora
- 1.5. Reyna, StephenDocumento21 pagine1.5. Reyna, StephenGomo RaNessuna valutazione finora
- CUSACK Otherkin TherianthropyDocumento16 pagineCUSACK Otherkin TherianthropyNafida TohirohNessuna valutazione finora
- University of Hawai'i Press Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Philosophy East and WestDocumento4 pagineUniversity of Hawai'i Press Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Philosophy East and WestJUAN CARLOS REYESNessuna valutazione finora
- Taylor - The Soul's Body and Its StatesDocumento8 pagineTaylor - The Soul's Body and Its StatesDavid HolzmerNessuna valutazione finora
- Wilson 1976Documento6 pagineWilson 1976Rafaella GaleanoNessuna valutazione finora
- Berghahn Books The Cambridge Journal of AnthropologyDocumento10 pagineBerghahn Books The Cambridge Journal of AnthropologyThiago TimNessuna valutazione finora
- Margaret Lock - Cultivating The BodyDocumento25 pagineMargaret Lock - Cultivating The BodyLuiz DuarteNessuna valutazione finora
- (Resenha) BOSKOVIC, Aleksandar. Johannes Fabian, Time and The Work of Anthropology - Critical Essays 1971-1991Documento4 pagine(Resenha) BOSKOVIC, Aleksandar. Johannes Fabian, Time and The Work of Anthropology - Critical Essays 1971-1991JRODNessuna valutazione finora
- Bessire & Bond 2014 Ontological Anthropology and The Deferral of Critique PDFDocumento17 pagineBessire & Bond 2014 Ontological Anthropology and The Deferral of Critique PDFnick reuterNessuna valutazione finora
- Roberts 2002Documento20 pagineRoberts 2002mariaconstanzavasqueNessuna valutazione finora
- Inconsciente en Levi-StraussDocumento29 pagineInconsciente en Levi-StraussjoakinenNessuna valutazione finora
- On The Nature of Being Human - TAYLORDocumento16 pagineOn The Nature of Being Human - TAYLORMateus MarinNessuna valutazione finora
- The Dawn of Everything A New History of HumanityDocumento5 pagineThe Dawn of Everything A New History of HumanityhiendangkyNessuna valutazione finora
- Super Report On Everything and BeyonfDocumento14 pagineSuper Report On Everything and BeyonfgruntyNessuna valutazione finora
- Jacques Berlinerblau, Toward A Sociology of Heresy, Orthodoxy, and DoxaDocumento26 pagineJacques Berlinerblau, Toward A Sociology of Heresy, Orthodoxy, and DoxaAnonymous hXQMt5Nessuna valutazione finora
- Evolutionary Psychology, Human Universals, and The Standard Social Science ModelDocumento14 pagineEvolutionary Psychology, Human Universals, and The Standard Social Science ModelkarinatyasNessuna valutazione finora
- Stocking - Franz Boas and The Culture Concept in Historical PerspectiveDocumento17 pagineStocking - Franz Boas and The Culture Concept in Historical PerspectiveShu-Yu Weng100% (1)
- Fuentes 2010 - On NatureDocumento10 pagineFuentes 2010 - On NatureTalita SeneNessuna valutazione finora
- Race and (The Study Of) EsotericismDocumento22 pagineRace and (The Study Of) EsotericismNicky KellyNessuna valutazione finora
- Roscoe 1995 The Perils of Positivism in Cultural AnthropologyDocumento14 pagineRoscoe 1995 The Perils of Positivism in Cultural AnthropologyBoris VelinesNessuna valutazione finora
- D. Schneider - The Meaning of IncestDocumento22 pagineD. Schneider - The Meaning of IncestNian PissolatiNessuna valutazione finora
- Philosophy of Language-SeminarDocumento3 paginePhilosophy of Language-SeminarInesNessuna valutazione finora
- Brown 2008 Cultural RelativismDocumento21 pagineBrown 2008 Cultural RelativismcoolkurllaNessuna valutazione finora
- Sanjay Seth - Once Was Blind But Now Can See Modernity and The Social SciencesDocumento16 pagineSanjay Seth - Once Was Blind But Now Can See Modernity and The Social SciencesQuelen GuedesNessuna valutazione finora
- CVTrueblood SR CWDocumento1 paginaCVTrueblood SR CWra monaNessuna valutazione finora
- Pet HolidyDocumento3 paginePet Holidyra monaNessuna valutazione finora
- BackupDocumento3 pagineBackupra monaNessuna valutazione finora
- English Teaching Handbook - BontilaDocumento39 pagineEnglish Teaching Handbook - BontilaEliza SorinaNessuna valutazione finora
- CVTrueblood SR CWDocumento1 paginaCVTrueblood SR CWra monaNessuna valutazione finora
- Raell PDFDocumento58 pagineRaell PDFopreamihNessuna valutazione finora
- Books 2 Check OnlineDocumento2 pagineBooks 2 Check Onlinera monaNessuna valutazione finora
- Movies To Be SeenDocumento3 pagineMovies To Be Seenra monaNessuna valutazione finora
- Links Tarski LimbajDocumento1 paginaLinks Tarski Limbajra monaNessuna valutazione finora
- The Innovations of The Twentieth-Century Novel: AbstractDocumento10 pagineThe Innovations of The Twentieth-Century Novel: Abstractsusom_iuliaNessuna valutazione finora
- TV Series List Mindhunter (2017-) The Gifted (2017 - Cosmos: A Spacetime Odyssey (2014-) Electric Dreams (2017-) Altered Carbon (2018-)Documento1 paginaTV Series List Mindhunter (2017-) The Gifted (2017 - Cosmos: A Spacetime Odyssey (2014-) Electric Dreams (2017-) Altered Carbon (2018-)ra monaNessuna valutazione finora
- 24224Documento5 pagine24224ra monaNessuna valutazione finora
- Teaching English Grammar: Mg. Evelyn Guillén ChávezDocumento43 pagineTeaching English Grammar: Mg. Evelyn Guillén Chávezra monaNessuna valutazione finora
- Complete-the-Second-Sentence-So-That-It-Has-a-Similar-Meaning-to-the-First-Sentence - Using-the-Word-Given AnswersDocumento6 pagineComplete-the-Second-Sentence-So-That-It-Has-a-Similar-Meaning-to-the-First-Sentence - Using-the-Word-Given AnswersMidNessuna valutazione finora
- Titularizare Writing StyleDocumento11 pagineTitularizare Writing Stylera monaNessuna valutazione finora
- Imaginary Homelands: Salman RushdieDocumento12 pagineImaginary Homelands: Salman Rushdiera monaNessuna valutazione finora
- NEVEN SESARDIC Making Sense of HeritabilDocumento8 pagineNEVEN SESARDIC Making Sense of Heritabilra monaNessuna valutazione finora
- 101 American English ProverbsDocumento1 pagina101 American English Proverbsra monaNessuna valutazione finora
- Variante Romana BacDocumento26 pagineVariante Romana Bacra mona0% (2)
- This Article Was Downloaded By: (Bowman, Paul) On: 15 April 2008Documento19 pagineThis Article Was Downloaded By: (Bowman, Paul) On: 15 April 2008bowman.webmailNessuna valutazione finora
- Literatura TitularizareDocumento1 paginaLiteratura Titularizarera monaNessuna valutazione finora
- Contextualize The Text From A Historical and Cultural Point of View - FFFF BunDocumento3 pagineContextualize The Text From A Historical and Cultural Point of View - FFFF BunAndreea100% (1)
- How To Build A Call Center From The Ground UpDocumento19 pagineHow To Build A Call Center From The Ground Upra monaNessuna valutazione finora
- Flyer Altitude Enterprise Recording en PDFDocumento2 pagineFlyer Altitude Enterprise Recording en PDFra monaNessuna valutazione finora
- Examexc PDFDocumento202 pagineExamexc PDFBudeanu AncaNessuna valutazione finora
- 2016 Spring LookbookDocumento46 pagine2016 Spring Lookbookra monaNessuna valutazione finora
- Useful LinksDocumento1 paginaUseful Linksra monaNessuna valutazione finora
- Copywriter 3.2016 EXTDocumento2 pagineCopywriter 3.2016 EXTra monaNessuna valutazione finora
- 10x Emails TemplatesDocumento8 pagine10x Emails TemplatesTestNessuna valutazione finora
- Conceptual Artist in Nigeria UNILAGDocumento13 pagineConceptual Artist in Nigeria UNILAGAdelekan FortuneNessuna valutazione finora
- Skincare Routine Order Cheat SheetDocumento10 pagineSkincare Routine Order Cheat SheetYel Salenga100% (3)
- Comparative Study On Serial and Parallel Manipulators - ReviewDocumento23 pagineComparative Study On Serial and Parallel Manipulators - ReviewShaik Himam SahebNessuna valutazione finora
- Islam and Civilization (Analysis Study On The History of Civilization in Islam) Muhammad Hifdil IslamDocumento18 pagineIslam and Civilization (Analysis Study On The History of Civilization in Islam) Muhammad Hifdil IslamLoveth KonniaNessuna valutazione finora
- Gondola CalculationDocumento6 pagineGondola CalculationBudi SusantoNessuna valutazione finora
- Carinthia Katalog DownloadDocumento16 pagineCarinthia Katalog DownloadOperator_010100% (2)
- Worksheet - 143760187HS-II, TUTORIAL ON CH-5Documento14 pagineWorksheet - 143760187HS-II, TUTORIAL ON CH-5A MusaverNessuna valutazione finora
- Clinical Reviews: The Management of Children With Gastroenteritis and Dehydration in The Emergency DepartmentDocumento13 pagineClinical Reviews: The Management of Children With Gastroenteritis and Dehydration in The Emergency DepartmentRina Dewi AnggraeniNessuna valutazione finora
- Ecall Vs NG EcallDocumento6 pagineEcall Vs NG EcallTrần Văn DũngNessuna valutazione finora
- Yoga SadhguruDocumento6 pagineYoga Sadhgurucosti.sorescuNessuna valutazione finora
- Liftchain (LC2A) Catalog AirHoistDocumento10 pagineLiftchain (LC2A) Catalog AirHoisteduardshark100% (1)
- Eco JetDocumento15 pagineEco JetJustin CoyNessuna valutazione finora
- W0L0XCF0866101640 (2006 Opel Corsa) PDFDocumento7 pagineW0L0XCF0866101640 (2006 Opel Corsa) PDFgianyNessuna valutazione finora
- Maintenance Páginas-509-580Documento72 pagineMaintenance Páginas-509-580Alexandra Gabriela Pacheco PrietoNessuna valutazione finora
- Solar Charge Controller: Solar Car Solar Home Solar Backpack Solar Boat Solar Street Light Solar Power GeneratorDocumento4 pagineSolar Charge Controller: Solar Car Solar Home Solar Backpack Solar Boat Solar Street Light Solar Power Generatorluis fernandoNessuna valutazione finora
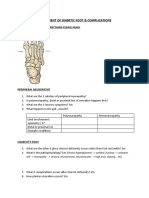
- Assessment of Diabetic FootDocumento7 pagineAssessment of Diabetic FootChathiya Banu KrishenanNessuna valutazione finora
- WeeklyDocumento8 pagineWeeklyivaldeztNessuna valutazione finora
- Tabla9 1Documento1 paginaTabla9 1everquinNessuna valutazione finora
- Ict 2120 Animation NC Ii Week 11 20 by Francis Isaac 1Documento14 pagineIct 2120 Animation NC Ii Week 11 20 by Francis Isaac 1Chiropractic Marketing NowNessuna valutazione finora
- Compiled LecsDocumento24 pagineCompiled LecsNur SetsuNessuna valutazione finora
- Regression Analysis Random Motors ProjectDocumento22 pagineRegression Analysis Random Motors ProjectPrateek AgrawalNessuna valutazione finora
- The Process: by Andy ZoppeltDocumento4 pagineThe Process: by Andy ZoppeltMark Stephen HuBertNessuna valutazione finora
- Maritime Management SystemsDocumento105 pagineMaritime Management SystemsAndika AntakaNessuna valutazione finora
- Birla MEEP Op ManualDocumento43 pagineBirla MEEP Op ManualAshok ChettiyarNessuna valutazione finora
- Aquaculture Scoop May IssueDocumento20 pagineAquaculture Scoop May IssueAquaculture ScoopNessuna valutazione finora
- Filipino Construction TermsDocumento6 pagineFilipino Construction TermsAdrian Perez75% (4)
- Synthesis Essay Coming To Grips With GenesisDocumento11 pagineSynthesis Essay Coming To Grips With Genesisapi-259381516Nessuna valutazione finora
- Medical GeneticsDocumento4 pagineMedical GeneticsCpopNessuna valutazione finora
- Dairy Products Theory XIIDocumento152 pagineDairy Products Theory XIIDskNessuna valutazione finora
- Book Index The Art of Heavy TransportDocumento6 pagineBook Index The Art of Heavy TransportHermon Pakpahan50% (2)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDa EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityValutazione: 4 su 5 stelle4/5 (24)
- Prisoners of Geography: Ten Maps That Explain Everything About the WorldDa EverandPrisoners of Geography: Ten Maps That Explain Everything About the WorldValutazione: 4.5 su 5 stelle4.5/5 (1145)
- Briefly Perfectly Human: Making an Authentic Life by Getting Real About the EndDa EverandBriefly Perfectly Human: Making an Authentic Life by Getting Real About the EndNessuna valutazione finora
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Da EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Valutazione: 4.5 su 5 stelle4.5/5 (110)
- 1177 B.C.: The Year Civilization Collapsed: Revised and UpdatedDa Everand1177 B.C.: The Year Civilization Collapsed: Revised and UpdatedValutazione: 4.5 su 5 stelle4.5/5 (111)
- Hey, Hun: Sales, Sisterhood, Supremacy, and the Other Lies Behind Multilevel MarketingDa EverandHey, Hun: Sales, Sisterhood, Supremacy, and the Other Lies Behind Multilevel MarketingValutazione: 4 su 5 stelle4/5 (103)
- The Wicked and the Willing: An F/F Gothic Horror Vampire NovelDa EverandThe Wicked and the Willing: An F/F Gothic Horror Vampire NovelValutazione: 4 su 5 stelle4/5 (21)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDa EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityValutazione: 4 su 5 stelle4/5 (3)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryDa EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryValutazione: 4 su 5 stelle4/5 (44)
- If I Did It: Confessions of the KillerDa EverandIf I Did It: Confessions of the KillerValutazione: 3 su 5 stelle3/5 (133)
- Selling the Dream: The Billion-Dollar Industry Bankrupting AmericansDa EverandSelling the Dream: The Billion-Dollar Industry Bankrupting AmericansValutazione: 4 su 5 stelle4/5 (17)
- Perfect Murder, Perfect Town: The Uncensored Story of the JonBenet Murder and the Grand Jury's Search for the TruthDa EverandPerfect Murder, Perfect Town: The Uncensored Story of the JonBenet Murder and the Grand Jury's Search for the TruthValutazione: 3.5 su 5 stelle3.5/5 (68)
- Troubled: A Memoir of Foster Care, Family, and Social ClassDa EverandTroubled: A Memoir of Foster Care, Family, and Social ClassValutazione: 4.5 su 5 stelle4.5/5 (26)
- The Exvangelicals: Loving, Living, and Leaving the White Evangelical ChurchDa EverandThe Exvangelicals: Loving, Living, and Leaving the White Evangelical ChurchValutazione: 4.5 su 5 stelle4.5/5 (13)
- Teaching to Transgress: Education as the Practice of FreedomDa EverandTeaching to Transgress: Education as the Practice of FreedomValutazione: 5 su 5 stelle5/5 (76)
- American Jezebel: The Uncommon Life of Anne Hutchinson, the Woman Who Defied the PuritansDa EverandAmerican Jezebel: The Uncommon Life of Anne Hutchinson, the Woman Who Defied the PuritansValutazione: 3.5 su 5 stelle3.5/5 (66)
- The Other Significant Others: Reimagining Life with Friendship at the CenterDa EverandThe Other Significant Others: Reimagining Life with Friendship at the CenterValutazione: 4 su 5 stelle4/5 (1)
- Body Language: Decode Human Behaviour and How to Analyze People with Persuasion Skills, NLP, Active Listening, Manipulation, and Mind Control Techniques to Read People Like a Book.Da EverandBody Language: Decode Human Behaviour and How to Analyze People with Persuasion Skills, NLP, Active Listening, Manipulation, and Mind Control Techniques to Read People Like a Book.Valutazione: 5 su 5 stelle5/5 (81)
- My Grandmother's Hands: Racialized Trauma and the Pathway to Mending Our Hearts and BodiesDa EverandMy Grandmother's Hands: Racialized Trauma and the Pathway to Mending Our Hearts and BodiesValutazione: 5 su 5 stelle5/5 (70)
- His Needs, Her Needs: Building a Marriage That LastsDa EverandHis Needs, Her Needs: Building a Marriage That LastsValutazione: 4.5 su 5 stelle4.5/5 (100)
- Disfigured: On Fairy Tales, Disability, and Making SpaceDa EverandDisfigured: On Fairy Tales, Disability, and Making SpaceValutazione: 4.5 su 5 stelle4.5/5 (396)