Potrebbero piacerti anche
- Aprender a ser salvajes: Cómo las culturas animales crían familias, crean belleza y consiguen la pazDa EverandAprender a ser salvajes: Cómo las culturas animales crían familias, crean belleza y consiguen la pazNessuna valutazione finora
- Presentación WallaceDocumento23 paginePresentación WallaceAdriana LopezNessuna valutazione finora
- Libro Origen de Las EspeciesDocumento67 pagineLibro Origen de Las EspeciesSerch MartinezNessuna valutazione finora
- El origen del hombre, la selección natural y la sexual.: Premium EbookDa EverandEl origen del hombre, la selección natural y la sexual.: Premium EbookValutazione: 4 su 5 stelle4/5 (1)
- Resumen Charles DarwinDocumento4 pagineResumen Charles DarwinSussi HerreraNessuna valutazione finora
- Teoría Evolutiva Charles DarwinDocumento7 pagineTeoría Evolutiva Charles DarwinAndreina GonzalezNessuna valutazione finora
- La Teoría de La EvoluciónDocumento5 pagineLa Teoría de La EvoluciónEdainBrocaTorresNessuna valutazione finora
- Biografia de Charles DarwinDocumento4 pagineBiografia de Charles DarwinÄdëlä Gönzälëz100% (5)
- 7 Teorias EvolutivasDocumento8 pagine7 Teorias EvolutivasYayaMilochoNessuna valutazione finora
- 3 Teorías de La Evolución.Documento35 pagine3 Teorías de La Evolución.Pedro JaraNessuna valutazione finora
- DarwinDocumento7 pagineDarwinEmilce EspíndolaNessuna valutazione finora
- Gomez David 4°1°Documento7 pagineGomez David 4°1°David GomezNessuna valutazione finora
- Resúmen Unidad UnoDocumento3 pagineResúmen Unidad Unomartuhoran1996Nessuna valutazione finora
- Teorias de La Evolucion Trabajo Final PDocumento19 pagineTeorias de La Evolucion Trabajo Final PWemy Wemy B FNessuna valutazione finora
- Evolucion Segun DarwinDocumento2 pagineEvolucion Segun Darwinmiavalent2023.scNessuna valutazione finora
- Tema 4Documento6 pagineTema 4bertin osborneNessuna valutazione finora
- Teoría de DarwinDocumento2 pagineTeoría de DarwinJesus Sanjuan MendezNessuna valutazione finora
- Biología IIDocumento29 pagineBiología IIzayra zhNessuna valutazione finora
- Teoria Karl PopperDocumento3 pagineTeoria Karl PopperAnthony CarrascoNessuna valutazione finora
- Origen y EvoluciónDocumento66 pagineOrigen y EvoluciónAlejadra BecerraNessuna valutazione finora
- Resumen Teoria de La Evolución Core UaiDocumento4 pagineResumen Teoria de La Evolución Core Uaimariajesus.losadarNessuna valutazione finora
- Resumen de La Teoría de DarwinDocumento6 pagineResumen de La Teoría de DarwinEsteban MaryoryNessuna valutazione finora
- Teorías de La Evolución y El Origen de Las EspeciesDocumento34 pagineTeorías de La Evolución y El Origen de Las EspeciesEvia Jhon100% (1)
- El Origen de La Vida MONOGRAFIA PARTE 2Documento27 pagineEl Origen de La Vida MONOGRAFIA PARTE 2David Salomón Flore OtuvoNessuna valutazione finora
- DarwinDocumento3 pagineDarwinAlexi LaiHoNessuna valutazione finora
- CMC-La Evolución (Tema 2)Documento11 pagineCMC-La Evolución (Tema 2)NachoAragónBéticoNessuna valutazione finora
- Ensayo Sobre La SelecciónDocumento3 pagineEnsayo Sobre La SelecciónAngeles Estrada GarciaNessuna valutazione finora
- Castro Nogueira & Toro Ibañez - Existe El AltruismoDocumento10 pagineCastro Nogueira & Toro Ibañez - Existe El Altruismojose.costajuniorNessuna valutazione finora
- Teoria de La EvolucionDocumento6 pagineTeoria de La EvolucionCamila BusquetNessuna valutazione finora
- Teorias de La EvolucionDocumento18 pagineTeorias de La EvolucionWemy Wemy B FNessuna valutazione finora
- Fijismo Vs Transformismo.Documento5 pagineFijismo Vs Transformismo.Miguel Benavides Acevedo0% (1)
- Seleccion NaturalDocumento8 pagineSeleccion NaturalRebeka GenesisNessuna valutazione finora
- Teorias de La EvoluciónDocumento20 pagineTeorias de La EvoluciónAracelli Claros BarreraNessuna valutazione finora
- La Teoría de DarwinDocumento15 pagineLa Teoría de DarwinPatty ChbNessuna valutazione finora
- Sva 04 - Teoría de La EvoluciónDocumento26 pagineSva 04 - Teoría de La EvoluciónARCINIEGA FLORES CHRISTOPHER BENJAMÍNNessuna valutazione finora
- Teorías EvolutivasDocumento4 pagineTeorías EvolutivasLeonardo MartinezNessuna valutazione finora
- Exposcion Teoria de DarwinDocumento5 pagineExposcion Teoria de DarwinMayelaNessuna valutazione finora
- Carlos DarwinDocumento2 pagineCarlos DarwinQpon gooNessuna valutazione finora
- Jean Batiste Lamarck La Diversidad Biológica Preguntas y RespuestasDocumento3 pagineJean Batiste Lamarck La Diversidad Biológica Preguntas y RespuestasCamCam VeuNessuna valutazione finora
- Clase 1 EvoluciónDocumento32 pagineClase 1 EvoluciónAntoniaNessuna valutazione finora
- Teoría Del Origen de La Especie Humana Y El Proceso de HumanizaciónDocumento7 pagineTeoría Del Origen de La Especie Humana Y El Proceso de Humanizaciónwww_yosoy50% (2)
- Alumnoscuestionario La Evolución de Las EspeciesDocumento2 pagineAlumnoscuestionario La Evolución de Las EspeciesRodrigo JohanNessuna valutazione finora
- Diapositivas Teoria de La Evolucion Power PointDocumento26 pagineDiapositivas Teoria de La Evolucion Power PointMARY FENIVER BORBON BARRETONessuna valutazione finora
- Resumen de Memoria y Aprendizaje (SELECCION NATURAL)Documento6 pagineResumen de Memoria y Aprendizaje (SELECCION NATURAL)psgagaNessuna valutazione finora
- Qué Es La EvoluciónDocumento3 pagineQué Es La EvoluciónWidman IxcoyNessuna valutazione finora
- Material de Lectura Seleccion NaturalDocumento3 pagineMaterial de Lectura Seleccion NaturalKathleen CastroNessuna valutazione finora
- Teorías Sobre El Origen de Las Especies, Biología-1° MedioDocumento8 pagineTeorías Sobre El Origen de Las Especies, Biología-1° MedioebellorNessuna valutazione finora
- Ensayo Sobre La SelecciónDocumento2 pagineEnsayo Sobre La SelecciónKelvin Antoni Benitres ValeroNessuna valutazione finora
- Ensayo Sobre La SelecciónDocumento2 pagineEnsayo Sobre La SelecciónKelvin Antoni Benitres ValeroNessuna valutazione finora
- Trabajo Biodiversidad y EvoluciónDocumento12 pagineTrabajo Biodiversidad y EvoluciónA Tempo Di EsperienzaNessuna valutazione finora
- Teoría Darwin-WallaceDocumento3 pagineTeoría Darwin-WallaceNoemí GilNessuna valutazione finora
- Ensayo Sobre El Origen de Las EspeciesDocumento4 pagineEnsayo Sobre El Origen de Las EspeciesCarlos100% (1)
- Macroevolucion I & II PTDocumento48 pagineMacroevolucion I & II PTjenniGNessuna valutazione finora
- Guia DarwinDocumento3 pagineGuia DarwinAndree MarquezNessuna valutazione finora
- Seleccion Natural BiologiaDocumento2 pagineSeleccion Natural BiologiaMontserrat GonzalezNessuna valutazione finora
- Evolución y BiodiversidadDocumento34 pagineEvolución y BiodiversidadCamila CastilloNessuna valutazione finora
- 4 Evolución y EspecieDocumento51 pagine4 Evolución y EspecieDaniela SepulvedaNessuna valutazione finora
- El FijismoDocumento4 pagineEl FijismoKaren BonisNessuna valutazione finora
- Teorias de La EvolucionDocumento10 pagineTeorias de La Evoluciondolly100% (2)
- Cuadratura de La Una Porción de Parábola PDFDocumento4 pagineCuadratura de La Una Porción de Parábola PDFLuchoNessuna valutazione finora
- Cómo Escribir Historia de La BiologíaDocumento16 pagineCómo Escribir Historia de La BiologíaLuchoNessuna valutazione finora
- Mendel para Enseñanza PDFDocumento8 pagineMendel para Enseñanza PDFLuchoNessuna valutazione finora
- Idealismo y MaterialismoDocumento2 pagineIdealismo y MaterialismoLuchoNessuna valutazione finora
- Abc Ciclo CelularDocumento2 pagineAbc Ciclo CelularDasio Buendia TaipeNessuna valutazione finora
- Guayacán: (Shirley) Escribir en El TableroDocumento3 pagineGuayacán: (Shirley) Escribir en El TableroWilson JaimesNessuna valutazione finora
- Guia#7 SextosDocumento2 pagineGuia#7 SextosLeoneRinconQuinteroNessuna valutazione finora
- Metarhizium AnisopliaeDocumento2 pagineMetarhizium AnisopliaeEstefaniaNessuna valutazione finora
- Creep FeedingDocumento10 pagineCreep FeedingRaul KimNessuna valutazione finora
- Informe 2 PaleoDocumento7 pagineInforme 2 PaleoEmersonNessuna valutazione finora
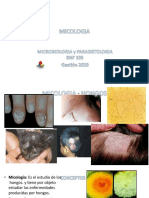
- MICOLOGIADocumento20 pagineMICOLOGIARashell rvNessuna valutazione finora
- Compendio Micro 2016Documento295 pagineCompendio Micro 2016Alfredo OyarzunNessuna valutazione finora
- Cuadro Comparativo Colénquima-Esclerénquima y ParénquimaDocumento1 paginaCuadro Comparativo Colénquima-Esclerénquima y ParénquimaLuis LealNessuna valutazione finora
- Principios y Criterios de BiocomercioDocumento20 paginePrincipios y Criterios de BiocomercioBioComercioPeruNessuna valutazione finora
- Características de Las Células (Erica)Documento2 pagineCaracterísticas de Las Células (Erica)xpress111100% (4)
- Primer Seminario MicrobiologíaDocumento21 paginePrimer Seminario MicrobiologíaAlejandra MoralesNessuna valutazione finora
- Reino AnimalDocumento21 pagineReino AnimalBryan Quenta100% (1)
- REACCION DE DIFERENTES GENOTIPOS DE TRIGO A LA PUDRICION DE ESPIGA CAUSADA POR Fusarium Graminearum PDFDocumento83 pagineREACCION DE DIFERENTES GENOTIPOS DE TRIGO A LA PUDRICION DE ESPIGA CAUSADA POR Fusarium Graminearum PDFCarlos Alberto Rosero DelgadoNessuna valutazione finora
- Protocolo CosmeticoDocumento4 pagineProtocolo CosmeticoItzel A. GuerreroNessuna valutazione finora
- INTA - Catalogo de Plantas para Techos Verdes PDFDocumento17 pagineINTA - Catalogo de Plantas para Techos Verdes PDFColo Colo100% (2)
- Karen - Alejandra - Fandiño - Grupo 18Documento6 pagineKaren - Alejandra - Fandiño - Grupo 18luis salazar osorioNessuna valutazione finora
- El BucareDocumento4 pagineEl BucareEduardo Hernandez100% (1)
- Diapositivas de VirusDocumento54 pagineDiapositivas de Virusgaxir80% (10)
- Microorganismos IndicadoresDocumento5 pagineMicroorganismos IndicadoresUriel RojasNessuna valutazione finora
- Tarea 1 Conducta AnimalDocumento7 pagineTarea 1 Conducta Animalrossy mejia100% (1)
- 11 Indicadores - EcológicosDocumento10 pagine11 Indicadores - Ecológicossteffany sanchezNessuna valutazione finora
- PERLITA ElSustratoPerfecto - Rev 2016Documento4 paginePERLITA ElSustratoPerfecto - Rev 2016J Sandro C-MNessuna valutazione finora
- Anelidos FinalDocumento10 pagineAnelidos FinalTeffy ColinaNessuna valutazione finora
- Triptico La Respiración de Los InvertebradosDocumento4 pagineTriptico La Respiración de Los InvertebradosPercy Mendoza67% (3)
- Selva MonzonicaDocumento5 pagineSelva MonzonicaaldoNessuna valutazione finora
- Clasificacion de Las HortalizasDocumento10 pagineClasificacion de Las HortalizasBerenisse Abigaly Aguilar TorresNessuna valutazione finora
- LOS VIRUS PresentaciónDocumento19 pagineLOS VIRUS PresentaciónJames SantiagoNessuna valutazione finora
- Ciencias Naturales Bloque 2 - 3Documento4 pagineCiencias Naturales Bloque 2 - 3Elia HernandezNessuna valutazione finora
- La Cadena AlimenticiaDocumento4 pagineLa Cadena AlimenticiaLuisNessuna valutazione finora