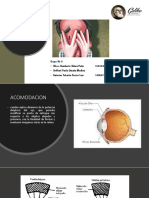
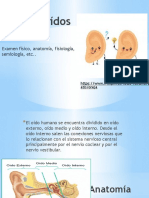
Potrebbero piacerti anche
- RESUMEN Vias VisualesDocumento5 pagineRESUMEN Vias VisualesValentina MuñozNessuna valutazione finora
- Función Nerviosa de La RetinaDocumento5 pagineFunción Nerviosa de La RetinaDAICY EDITH LEDEZMA PEREZNessuna valutazione finora
- Tracto ÓpticoDocumento14 pagineTracto ÓpticoYave Set Rosas Rodriguez0% (1)
- 5.3 Fisiología de Entrada de La Luz y Reflejos PupilaresDocumento41 pagine5.3 Fisiología de Entrada de La Luz y Reflejos PupilaresOscar Padilla Navarro100% (1)
- Función Receptora y Neural de La RetinaDocumento8 pagineFunción Receptora y Neural de La RetinaKaren LopezNessuna valutazione finora
- Vía Visual Óptica UnamDocumento3 pagineVía Visual Óptica UnamEvans Fernando EscobarNessuna valutazione finora
- El OjoDocumento4 pagineEl OjotuyopsonNessuna valutazione finora
- Embriogenesis Del Ojo 2014Documento52 pagineEmbriogenesis Del Ojo 2014WalterQunoqYanceOw100% (1)
- Corteza VisualDocumento15 pagineCorteza VisualDouglasNessuna valutazione finora
- Fisiologia Del OjoDocumento87 pagineFisiologia Del OjoALVARONessuna valutazione finora
- Neurofisiologia Central de La Vision PDFDocumento21 pagineNeurofisiologia Central de La Vision PDFDu LoredNessuna valutazione finora
- 8.2 Fisiología y Bioquímica Celular Del EPR (PEDRO)Documento26 pagine8.2 Fisiología y Bioquímica Celular Del EPR (PEDRO)Oscar Padilla Navarro100% (1)
- Sistemas SensorialesDocumento2 pagineSistemas SensorialesSamira100% (2)
- Camara Anterior Del OjoDocumento2 pagineCamara Anterior Del OjoPedro Santana LlamasNessuna valutazione finora
- CoroidesDocumento4 pagineCoroidessandra juliana nuñez lambrañoNessuna valutazione finora
- Óptica Y Ondas (Fisica)Documento26 pagineÓptica Y Ondas (Fisica)Cristian Villegas100% (1)
- Optica FisiologicaDocumento13 pagineOptica FisiologicaRubén León SuarezNessuna valutazione finora
- El Sistema VisualDocumento11 pagineEl Sistema VisualNorma Beatriz GonzalezNessuna valutazione finora
- 10 Capas de La RetinaDocumento2 pagine10 Capas de La RetinaNaeli ChampoNessuna valutazione finora
- FOTORRECEPCIÓNDocumento61 pagineFOTORRECEPCIÓNGustavoNessuna valutazione finora
- Neurofisiología de La VisiónDocumento8 pagineNeurofisiología de La VisiónRondon JsefranNessuna valutazione finora
- Oft Clase 1 GeneralidadesDocumento24 pagineOft Clase 1 GeneralidadesPaloma Mavier Marmolejos Mejia100% (1)
- AnatomiaDocumento12 pagineAnatomiaPaola Ortega FrancoNessuna valutazione finora
- Desarrollo Cronologico de Las Estructuras OcularesDocumento21 pagineDesarrollo Cronologico de Las Estructuras OcularesjoseNessuna valutazione finora
- Esclera, Iris y Pupila PAODocumento41 pagineEsclera, Iris y Pupila PAOjesusNessuna valutazione finora
- Fisiologia OcularDocumento23 pagineFisiologia OcularIsamar Pidiachi100% (1)
- Barreras Ha y HRDocumento22 pagineBarreras Ha y HROscar Padilla NavarroNessuna valutazione finora
- Tecnica de Iluminacion DirectaDocumento45 pagineTecnica de Iluminacion DirectaLuis Antonio CastañonNessuna valutazione finora
- Vias de La VisionDocumento2 pagineVias de La VisionRomario Paz100% (1)
- UVEA1Documento6 pagineUVEA1Argenis Parra OviedoNessuna valutazione finora
- Patologia OcularDocumento14 paginePatologia OcularShyannaNessuna valutazione finora
- GENESER HISTOLOGIA CAPITULO 24 El Ojo PDFDocumento42 pagineGENESER HISTOLOGIA CAPITULO 24 El Ojo PDFjoaquin rothNessuna valutazione finora
- BAJA VISION ExposicionDocumento40 pagineBAJA VISION Exposicionmanuel villavicencioNessuna valutazione finora
- Ambliopía Clase 6-08Documento7 pagineAmbliopía Clase 6-08May Suicidaal QuinnNessuna valutazione finora
- Estudio de La Diplopía y CoordimetriaDocumento10 pagineEstudio de La Diplopía y CoordimetriaAlondra Verdugo GómezNessuna valutazione finora
- 09 El Cristalino PDFDocumento151 pagine09 El Cristalino PDFEnmanuel Nazzar JMNessuna valutazione finora
- Oculomotor Testing PDFDocumento24 pagineOculomotor Testing PDFdavid peñaNessuna valutazione finora
- Ambliopía Ok OkDocumento30 pagineAmbliopía Ok OkAngie LadinoNessuna valutazione finora
- La Ambliopia y Su Tratamiento Neuro SensorialDocumento25 pagineLa Ambliopia y Su Tratamiento Neuro SensorialLuz Maria Valverde RamosNessuna valutazione finora
- Master en Optometria ClinicaDocumento30 pagineMaster en Optometria ClinicaAndrea Carolina VeraNessuna valutazione finora
- Modelos ExplicativosDocumento70 pagineModelos ExplicativosMaria AlejandraNessuna valutazione finora
- Historia Se Los Lentes de ContactoDocumento2 pagineHistoria Se Los Lentes de ContactoJûâň Đīēğø Jř González NavarroNessuna valutazione finora
- ESQUIASDocumento3 pagineESQUIASValentine Fritz100% (1)
- Anexos OcularesDocumento4 pagineAnexos Ocularessarita escobarNessuna valutazione finora
- Anatomia Globo Ocular1146Documento16 pagineAnatomia Globo Ocular1146lialeonorNessuna valutazione finora
- Vía VisualDocumento36 pagineVía Visualamy100% (1)
- 1 Anatomia y Fisiologia de Los Parpados Gonzalez Arias LucyDocumento5 pagine1 Anatomia y Fisiologia de Los Parpados Gonzalez Arias LucyMaría Patricia FereiraNessuna valutazione finora
- El Optico Frente A La Vision SubnormalDocumento18 pagineEl Optico Frente A La Vision SubnormalMonica Laura Sanchez100% (1)
- Tratamiento de La Ambliopía Basado en La Evidencia Científica Marzo 2020 PDFDocumento25 pagineTratamiento de La Ambliopía Basado en La Evidencia Científica Marzo 2020 PDFDavidNessuna valutazione finora
- Anexos Oculares Trabajo ExpoDocumento26 pagineAnexos Oculares Trabajo ExpoSantkiz Stk75% (4)
- Embriología OcularDocumento6 pagineEmbriología OcularMaría Ángeles NeiraNessuna valutazione finora
- Capitulo6 PDFDocumento110 pagineCapitulo6 PDFpatrisaintNessuna valutazione finora
- Pelicula LagrimalDocumento1 paginaPelicula LagrimalMitzy NavarreteNessuna valutazione finora
- Acomodación y PresbiciaDocumento54 pagineAcomodación y PresbiciaResidentes 2020Nessuna valutazione finora
- Óptica de FotorreceptoresDocumento22 pagineÓptica de FotorreceptoresNaNa SaldañaNessuna valutazione finora
- La Distribución de Conos y Bastones en La RetinaDocumento2 pagineLa Distribución de Conos y Bastones en La RetinaCarlos ceballosNessuna valutazione finora
- Anatomia y Fisiologia Del Globo Ocular EnsayoDocumento1 paginaAnatomia y Fisiologia Del Globo Ocular EnsayoManuel MirandaNessuna valutazione finora
- Seminario de Retinopatia Diabetica DapiDocumento26 pagineSeminario de Retinopatia Diabetica DapiRey FelixNessuna valutazione finora
- Vision Binocular Alvarez TapiasDocumento12 pagineVision Binocular Alvarez TapiasFranklin KerberNessuna valutazione finora
- Areas Del Cerebro 1Documento4 pagineAreas Del Cerebro 1ADRIANA GARCIANessuna valutazione finora
- Codigo Etica Guia IvDocumento70 pagineCodigo Etica Guia IvIrma Flores100% (1)
- Gerente GeneralDocumento3 pagineGerente GeneralIrma FloresNessuna valutazione finora
- Gerente GeneralDocumento3 pagineGerente GeneralIrma FloresNessuna valutazione finora
- Código de Ética Profesional Del Contador PúblicoDocumento16 pagineCódigo de Ética Profesional Del Contador PúblicoJosé CastañedaNessuna valutazione finora
- 11 Producto UGDocumento15 pagine11 Producto UGIrma FloresNessuna valutazione finora
- Codigo Etica Guia IiiDocumento18 pagineCodigo Etica Guia IiiIrma FloresNessuna valutazione finora
- 12 PrecioDocumento18 pagine12 PrecioIrma FloresNessuna valutazione finora
- Tesis55 PDFDocumento224 pagineTesis55 PDFPaulina WörnerNessuna valutazione finora
- Presentacion Alteraciones SensorialesDocumento22 paginePresentacion Alteraciones SensorialesIrma FloresNessuna valutazione finora
- 14 PromocionDocumento19 pagine14 PromocionIrma FloresNessuna valutazione finora
- 6th Central Pay Commission Salary CalculatorDocumento15 pagine6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- 13 PlazaDocumento10 pagine13 PlazaIrma FloresNessuna valutazione finora
- 13 PlazaDocumento10 pagine13 PlazaIrma FloresNessuna valutazione finora
- GLOSARIO RIGIDOS ResueltosDocumento2 pagineGLOSARIO RIGIDOS ResueltosIrma Flores76% (17)
- Bela Planodeplanta OPTICA 2 PDFDocumento1 paginaBela Planodeplanta OPTICA 2 PDFIrma FloresNessuna valutazione finora
- Optom. Terap. 6Documento27 pagineOptom. Terap. 6Irma FloresNessuna valutazione finora
- Forias y TropiasDocumento36 pagineForias y TropiasIrma Flores75% (4)
- Opt. Terap. 3Documento24 pagineOpt. Terap. 3Irma FloresNessuna valutazione finora
- Optom. Terap. 5Documento21 pagineOptom. Terap. 5Irma Flores100% (1)
- Como Hacer Microscopio CompuestoDocumento26 pagineComo Hacer Microscopio CompuestoIrma FloresNessuna valutazione finora
- Optm. Terap. 4Documento35 pagineOptm. Terap. 4Irma FloresNessuna valutazione finora
- Relacion Acomodoci N ConvergenciaDocumento25 pagineRelacion Acomodoci N ConvergenciaIrma FloresNessuna valutazione finora
- Motilidad OcularDocumento33 pagineMotilidad OcularIrma FloresNessuna valutazione finora
- Test Refractivo 4Documento5 pagineTest Refractivo 4Irma Flores100% (1)
- Practica Soo 1Documento72 paginePractica Soo 1Irma Flores100% (1)
- Primera Ley de NewtonDocumento1 paginaPrimera Ley de NewtonIrma FloresNessuna valutazione finora
- Acta CertificadaDocumento1 paginaActa CertificadaIrma FloresNessuna valutazione finora
- Mujer TrabajadoraDocumento3 pagineMujer TrabajadoraIrma FloresNessuna valutazione finora
- Trastornos de La VisiónDocumento2 pagineTrastornos de La VisiónIRENE SANTOS LAVILLANessuna valutazione finora
- Practica ..Documento5 paginePractica ..GIAN CARLO MAMANI GILES.Nessuna valutazione finora
- Consentimiento Informado para Cirugia de Vitreo y Retina PDFDocumento4 pagineConsentimiento Informado para Cirugia de Vitreo y Retina PDFveronica lizethNessuna valutazione finora
- Presentaciónsensacion y Percepcion.Documento24 paginePresentaciónsensacion y Percepcion.david ValleNessuna valutazione finora
- Full Modelos de PuertasDocumento32 pagineFull Modelos de PuertasGiovanny SanchezNessuna valutazione finora
- Hermann SnellenDocumento9 pagineHermann SnellenRubi Lopez EspadaNessuna valutazione finora
- Color eDocumento124 pagineColor eAlejandra Ramírez VázquezNessuna valutazione finora
- Agudeza VisualDocumento1 paginaAgudeza VisualborisNessuna valutazione finora
- Los Oídos SemiologiaDocumento43 pagineLos Oídos SemiologiaLuisarlin DiazNessuna valutazione finora
- Patologias Del CristalinoDocumento9 paginePatologias Del CristalinojoismarNessuna valutazione finora
- Guia Glaucoma 2019 FinalDocumento49 pagineGuia Glaucoma 2019 FinalNidia M. Quispe Rodríguez100% (2)
- Bonebridge: Tecnología de Vanguardia en Sistemas AuditivosDocumento20 pagineBonebridge: Tecnología de Vanguardia en Sistemas AuditivosmarciaNessuna valutazione finora
- PDF Vision Binocular Diagnostico y Tratamiento 01 CompressDocumento15 paginePDF Vision Binocular Diagnostico y Tratamiento 01 CompressMilton EstradaNessuna valutazione finora
- OptometriaDocumento4 pagineOptometriamaria sotoNessuna valutazione finora
- Quiz 2 - Semana 7 - RA - SEGUNDO BLOQUE-SENSACION Y PERCEPCION - (GRUPO1) Watsha.1Documento6 pagineQuiz 2 - Semana 7 - RA - SEGUNDO BLOQUE-SENSACION Y PERCEPCION - (GRUPO1) Watsha.1Katherine ToroNessuna valutazione finora
- Trabajo 1 Sistemas SensorialesDocumento15 pagineTrabajo 1 Sistemas SensorialesnitacitoNessuna valutazione finora
- Informe 2 - Angie FloresDocumento12 pagineInforme 2 - Angie FloresGugatienda DosNessuna valutazione finora
- Los Órganos de Los SentidosDocumento13 pagineLos Órganos de Los Sentidosdolores ecijaNessuna valutazione finora
- Sensación y PercepciónDocumento20 pagineSensación y PercepciónMaría Agustina MercadoNessuna valutazione finora
- Factores Protectores para Disminuir La Progresión de Miopía en Los NiñosDocumento15 pagineFactores Protectores para Disminuir La Progresión de Miopía en Los NiñosAlejandraOzunaSolanoNessuna valutazione finora
- El Sistema Sensorial y Sus Alteraciones PsicológicasDocumento9 pagineEl Sistema Sensorial y Sus Alteraciones PsicológicasMarlen NinaNessuna valutazione finora
- Libro - Manual - Teorico - Practico - en - Oftalmologia 2Documento111 pagineLibro - Manual - Teorico - Practico - en - Oftalmologia 2ERIKANessuna valutazione finora
- Organo VestibulococlearDocumento2 pagineOrgano VestibulococlearJavier WafflesNessuna valutazione finora
- Anatomia OidoDocumento21 pagineAnatomia Oidoapi-382656780% (5)
- Histologia Biologia Celular Fortoul Castell PDFDocumento306 pagineHistologia Biologia Celular Fortoul Castell PDFValentina Vásquez Arteaga100% (7)
- Astigmatismo y PresbiciaDocumento8 pagineAstigmatismo y PresbiciaVictor BarrezuetaNessuna valutazione finora
- Clase - DG AudiológicoDocumento28 pagineClase - DG AudiológicoDiegoNessuna valutazione finora
- Activacion 6 de Agosto Al 25Documento24 pagineActivacion 6 de Agosto Al 25Santiago AgudeloNessuna valutazione finora
- Los Sentidos HumanosDocumento15 pagineLos Sentidos HumanosVikttor PatatuchyNessuna valutazione finora
- Acustica (Ambiental - Arquitectonica)Documento92 pagineAcustica (Ambiental - Arquitectonica)EnriqueNessuna valutazione finora