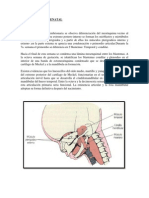
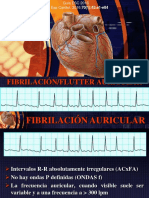
Potrebbero piacerti anche
- Desarrollo Del PeriodontoDocumento16 pagineDesarrollo Del PeriodontoNinja Nikitay50% (2)
- Genes HoxDocumento12 pagineGenes HoxAndrea Cardenas FrancoNessuna valutazione finora
- 1.presentación Modelo Cero Accidentes 2013Documento14 pagine1.presentación Modelo Cero Accidentes 2013Stela MosqueraNessuna valutazione finora
- Tecnicas y Estrategias Metodologicas para El Desarrollo de Habilidades Cognitivas y SocialesDocumento12 pagineTecnicas y Estrategias Metodologicas para El Desarrollo de Habilidades Cognitivas y SocialesJhofre Chavez Pastor100% (3)
- Técnicas CriptográficasDocumento12 pagineTécnicas CriptográficasBubosqui Buba0% (1)
- Aporte de Las Culturas Antiguas en La OdontologíaDocumento35 pagineAporte de Las Culturas Antiguas en La OdontologíaAnabel Vinueza100% (1)
- Histologia, Cemento, Periodon, Hueso 1Documento68 pagineHistologia, Cemento, Periodon, Hueso 1Leonardo Manuel VillegasNessuna valutazione finora
- Microbiología BucalDocumento14 pagineMicrobiología BucalJ Daniel LlanosNessuna valutazione finora
- Desarrollo Del CATMDocumento6 pagineDesarrollo Del CATMMelany Del PilarNessuna valutazione finora
- Guía de Póster de Adhesión en OdontologíaDocumento3 pagineGuía de Póster de Adhesión en OdontologíamarcellNessuna valutazione finora
- CATM Desarrollo PrenatalDocumento7 pagineCATM Desarrollo PrenatalKarina Marly SantanaNessuna valutazione finora
- Resumen BiofilmDocumento5 pagineResumen BiofilmCitlalli RguezNessuna valutazione finora
- Clases de Periodoncia Ro2 Michel SuarezDocumento29 pagineClases de Periodoncia Ro2 Michel SuarezDANIELA CALDERONNessuna valutazione finora
- El Esmalte DentalDocumento22 pagineEl Esmalte DentalAndrea Ramon nuñezNessuna valutazione finora
- Generalidades de Resinas CompuestasDocumento9 pagineGeneralidades de Resinas CompuestasAhre WanakoNessuna valutazione finora
- Examenes Complementarios en OdontologiaDocumento18 pagineExamenes Complementarios en OdontologiaMiguel HernändezNessuna valutazione finora
- Amelogenesis GRAN MARISCAL DE AYACUCHODocumento4 pagineAmelogenesis GRAN MARISCAL DE AYACUCHODoomVoiceNessuna valutazione finora
- Opiodes 2023Documento12 pagineOpiodes 2023ERIN CHARLIZE ALECIO MARTINEZNessuna valutazione finora
- InfografíaDocumento8 pagineInfografíaMichelle Maldonado100% (1)
- Mucosa BucalDocumento23 pagineMucosa BucalDeniss Rios Arostegui100% (1)
- Histología Del CementoDocumento9 pagineHistología Del CementoMarco QuispeNessuna valutazione finora
- Clase 4. - Pelicula Adquirida.Documento20 pagineClase 4. - Pelicula Adquirida.Josefa VenegasNessuna valutazione finora
- ENVEJECIMIENTO - Cambios Bioquímicos - ZentenoDocumento7 pagineENVEJECIMIENTO - Cambios Bioquímicos - ZentenoCarlos Albert Krus100% (1)
- Eje Temático 1 Generalidades Acerca de Odontología RestauradoraDocumento39 pagineEje Temático 1 Generalidades Acerca de Odontología RestauradoraLuis TR100% (1)
- Amelogenesis FinalDocumento34 pagineAmelogenesis FinalMario MedinaNessuna valutazione finora
- Complejo Dentino-PulparDocumento39 pagineComplejo Dentino-PulparJuan BernalNessuna valutazione finora
- 14 Antimicrobianos de Uso OdontológicoDocumento115 pagine14 Antimicrobianos de Uso OdontológicoMichelle Zazueta Rivera0% (1)
- Condiciones No Patológicas de La Cavidad BucalDocumento6 pagineCondiciones No Patológicas de La Cavidad BucalJoseLuisEspinozaBeltranNessuna valutazione finora
- Complejo Dentino PulparDocumento69 pagineComplejo Dentino PulparMary Oviedo BuilesNessuna valutazione finora
- Primer Premolar SuperiorDocumento4 paginePrimer Premolar SuperiorSidney Del Carmen Sumner CaballeroNessuna valutazione finora
- Ligamento PeriodontalDocumento6 pagineLigamento PeriodontalLeidi Cuenca100% (1)
- Materiales Elastoméricos I y IIDocumento12 pagineMateriales Elastoméricos I y IILidia Muñoz CastroNessuna valutazione finora
- Registros InteroclusalesDocumento18 pagineRegistros InteroclusalesSarai MoraNessuna valutazione finora
- 7 - UNIDAD Tejidos PeriodontalesDocumento120 pagine7 - UNIDAD Tejidos PeriodontalesCesia Carolina AvilaNessuna valutazione finora
- Primero Segundo y Tercer Molar SuperiorDocumento24 paginePrimero Segundo y Tercer Molar SuperiorRogerNessuna valutazione finora
- MacrolidosDocumento14 pagineMacrolidosKaren EsquivelNessuna valutazione finora
- Oclusion Terapeutica Desde Las Escuelas de Oclusion Ala OBE PDFDocumento6 pagineOclusion Terapeutica Desde Las Escuelas de Oclusion Ala OBE PDFPaulChillogalloNessuna valutazione finora
- Teorías de La Caries DentalDocumento11 pagineTeorías de La Caries Dentalmariana coronaNessuna valutazione finora
- Oclusión - Mecánica Del Movimiento Mandibular. (Autoguardado)Documento20 pagineOclusión - Mecánica Del Movimiento Mandibular. (Autoguardado)Mafer AraujoNessuna valutazione finora
- Teorías CariologíaDocumento7 pagineTeorías CariologíaJulieta FHNessuna valutazione finora
- Mucosa OralDocumento10 pagineMucosa OralAna María AramayoNessuna valutazione finora
- Anestesicos Locales en Odontologia - Leon, Miguel EvelioDocumento6 pagineAnestesicos Locales en Odontologia - Leon, Miguel EvelioKevin Emilio VargasNessuna valutazione finora
- Alteraciones PulparesDocumento26 pagineAlteraciones PulparesAnthony GamarraNessuna valutazione finora
- Bioquimica de La Caries DentalDocumento11 pagineBioquimica de La Caries DentalSachell ZavalaNessuna valutazione finora
- Estadio de CampanaDocumento4 pagineEstadio de CampanaPauNessuna valutazione finora
- Caries DentalDocumento5 pagineCaries DentalAnabel Silva RiosNessuna valutazione finora
- Icdas Clasificacion de Lesiones CariosasDocumento2 pagineIcdas Clasificacion de Lesiones CariosasJune Marilith Martinez CastellanosNessuna valutazione finora
- Actividad 1 Biologia Del Complejo Dentino PulparDocumento6 pagineActividad 1 Biologia Del Complejo Dentino PulparKarla TorresNessuna valutazione finora
- Evolucion Morfofuncional de La Denticion HumanaDocumento44 pagineEvolucion Morfofuncional de La Denticion HumanaKaory Carrasco100% (1)
- Alteraciones Pulpares y PeriapicalesDocumento6 pagineAlteraciones Pulpares y PeriapicalesMorales Gómez Angel TadeoNessuna valutazione finora
- Materiales Hemostáticos en Cirugía OralDocumento5 pagineMateriales Hemostáticos en Cirugía OralApque Patrick100% (2)
- Patologia HematicaDocumento25 paginePatologia HematicaOctavio CansignoNessuna valutazione finora
- Papiloma PlanoDocumento19 paginePapiloma PlanoJustin KelleyNessuna valutazione finora
- Pulpa Dental ExposicionDocumento39 paginePulpa Dental ExposicionKamilo BermudezNessuna valutazione finora
- Bioquimica de La Pulpa Dental GrupoDocumento26 pagineBioquimica de La Pulpa Dental GrupoNicole Vidalón PalominoNessuna valutazione finora
- La DentinaDocumento8 pagineLa DentinaVelez StalynNessuna valutazione finora
- Patobiologia PeriapicalDocumento62 paginePatobiologia PeriapicalNataliLazo100% (1)
- Ensayo Prevotella Intermedia - Jourgen Galvis HerreraDocumento5 pagineEnsayo Prevotella Intermedia - Jourgen Galvis HerreraJourgenNessuna valutazione finora
- Estudio Químico Cuántico de La Adsorción de Tiofeno y Tiofenos Metilados en Zeolita ÁcidaDocumento6 pagineEstudio Químico Cuántico de La Adsorción de Tiofeno y Tiofenos Metilados en Zeolita ÁcidaNacho MezaNessuna valutazione finora
- N Alquilacion de Aminas AromaticasDocumento10 pagineN Alquilacion de Aminas AromaticasMaritza Ximena Alonso MartinezNessuna valutazione finora
- Cromatografía Fina y de Columna.Documento4 pagineCromatografía Fina y de Columna.Abel de Jesús RiveraNessuna valutazione finora
- Indicadores de EficienciaDocumento5 pagineIndicadores de EficienciaAlvaro Portal LozanoNessuna valutazione finora
- LARINGOTRAQUEITISDocumento2 pagineLARINGOTRAQUEITISAlvaro Portal LozanoNessuna valutazione finora
- LEISHMANIASISDocumento3 pagineLEISHMANIASISAlvaro Portal LozanoNessuna valutazione finora
- n2 098 105 JuanMolinaDocumento9 paginen2 098 105 JuanMolinaEdgar LoezaNessuna valutazione finora
- Impacto de Las Enfermedades DiarreicasDocumento9 pagineImpacto de Las Enfermedades DiarreicasStefani RSNessuna valutazione finora
- Articulo para ExposicionDocumento14 pagineArticulo para Exposiciontuntun75Nessuna valutazione finora
- Diarrea AgudaDocumento40 pagineDiarrea AgudaJulioce WaldoNessuna valutazione finora
- El Gobierno de La UniversidadDocumento11 pagineEl Gobierno de La UniversidadAlvaro Portal LozanoNessuna valutazione finora
- Enfermedades Pediátricas DiscapacitantesDocumento7 pagineEnfermedades Pediátricas DiscapacitantesAlvaro Portal LozanoNessuna valutazione finora
- Prevencion y Manejo de Condiciones Secundarias de Salud en Personas Con DiscapacidadDocumento169 paginePrevencion y Manejo de Condiciones Secundarias de Salud en Personas Con DiscapacidadAlvaro Portal LozanoNessuna valutazione finora
- Naturaleza de La UniversidadDocumento12 pagineNaturaleza de La UniversidadAlvaro Portal LozanoNessuna valutazione finora
- Sociedad de Conocimiento y Problematica de La UniversidadDocumento7 pagineSociedad de Conocimiento y Problematica de La UniversidadAlvaro Portal LozanoNessuna valutazione finora
- Prevencion y Control Del CancerDocumento339 paginePrevencion y Control Del CancerAlvaro Portal LozanoNessuna valutazione finora
- MarcapasoDocumento35 pagineMarcapasoAlvaro Portal LozanoNessuna valutazione finora
- Reforma de CórdobaDocumento5 pagineReforma de CórdobaAlvaro Portal LozanoNessuna valutazione finora
- CORTES DE CEREBRO. Forlizzi, Pró. Universidad de Buenos AiresDocumento36 pagineCORTES DE CEREBRO. Forlizzi, Pró. Universidad de Buenos AiresAdriana Saavedra Martinez100% (2)
- INYECTABLESDocumento51 pagineINYECTABLESAlvaro Portal Lozano100% (1)
- Enfermedades Pediátricas DiscapacitantesDocumento7 pagineEnfermedades Pediátricas DiscapacitantesAlvaro Portal LozanoNessuna valutazione finora
- Anatomiayfisiologiadeviasrespiratorias 150311215232 Conversion Gate01Documento24 pagineAnatomiayfisiologiadeviasrespiratorias 150311215232 Conversion Gate01DIEGO ALEJANDRO MORALES CAYCEDONessuna valutazione finora
- Fibrilación AuricularDocumento37 pagineFibrilación AuricularAlvaro Portal LozanoNessuna valutazione finora
- Anatomia Presentacion2 161112113853Documento35 pagineAnatomia Presentacion2 161112113853Alvaro Portal LozanoNessuna valutazione finora
- Bocio y Enfermedad Tiroidea Nodular PDFDocumento33 pagineBocio y Enfermedad Tiroidea Nodular PDFAlvaro Portal LozanoNessuna valutazione finora
- Caso Clínico MielomaDocumento26 pagineCaso Clínico MielomaAlvaro Portal LozanoNessuna valutazione finora
- Trabajo de Monografia .EtsDocumento32 pagineTrabajo de Monografia .EtsAlvaro Portal LozanoNessuna valutazione finora
- Cómo Elaborar Una MonografíaDocumento24 pagineCómo Elaborar Una MonografíaAlvaro Portal Lozano100% (1)
- Informe de Disección Cabeza y CuelloDocumento27 pagineInforme de Disección Cabeza y CuelloAlvaro Portal LozanoNessuna valutazione finora
- Seminario 08 - Metabolismo Del CortisolDocumento29 pagineSeminario 08 - Metabolismo Del CortisolLuisAlfredoAlvaradoZavaletaNessuna valutazione finora
- Monografía EmbriologíaDocumento54 pagineMonografía EmbriologíaAlvaro Portal LozanoNessuna valutazione finora
- FORODocumento2 pagineFOROIlse CastilloNessuna valutazione finora
- Evolucion Del Pensamiento Filosofico Viii PDFDocumento6 pagineEvolucion Del Pensamiento Filosofico Viii PDFMarino SalcedoNessuna valutazione finora
- Antecedentes de La IA en La Industria 4.0Documento3 pagineAntecedentes de La IA en La Industria 4.0daniel sebastian saenz rangelNessuna valutazione finora
- Autoevaluacion 1Documento2 pagineAutoevaluacion 1Juan Heladio Sandoval SanchezNessuna valutazione finora
- Estadistica ResumenDocumento5 pagineEstadistica ResumennanobiusNessuna valutazione finora
- Llenado de Fua Salud Mental - 2019 Diresa - TumbesDocumento11 pagineLlenado de Fua Salud Mental - 2019 Diresa - TumbesCarlos CoronadoNessuna valutazione finora
- Las Abejas y Los ArquitectosDocumento7 pagineLas Abejas y Los ArquitectosjhordanNessuna valutazione finora
- Esquema Metadatos Gestión Documento ElectrónicoDocumento55 pagineEsquema Metadatos Gestión Documento ElectrónicopaolaespNessuna valutazione finora
- Plan de Trabajo Del Taller de TeatroDocumento6 paginePlan de Trabajo Del Taller de TeatroAuxiliares EducaciónunidosalsutepNessuna valutazione finora
- Antecedentes y Resumen de FisiocraciaDocumento3 pagineAntecedentes y Resumen de FisiocraciaMaria René RañaNessuna valutazione finora
- SEPARATA 2. Editada DENDRO 07.09.13Documento21 pagineSEPARATA 2. Editada DENDRO 07.09.13Anonymous 2HShWKOINessuna valutazione finora
- Semiología de La Patología Vascular en Los HemisferiosDocumento74 pagineSemiología de La Patología Vascular en Los HemisferiosCarlos VaronNessuna valutazione finora
- Arte y Cultura.Documento9 pagineArte y Cultura.JUDITSIRRYNessuna valutazione finora
- Jose Salazar Inglés 4C B2Documento2 pagineJose Salazar Inglés 4C B2jose miguel salazar sosaNessuna valutazione finora
- La Universidad de Ayer y HoyDocumento44 pagineLa Universidad de Ayer y HoyEmmanuel GiotNessuna valutazione finora
- Contaminacion AcusticaDocumento4 pagineContaminacion AcusticaDiego Chiroque RamirezNessuna valutazione finora
- Símbolos Patrios Del Estado AnzoáteguiDocumento9 pagineSímbolos Patrios Del Estado AnzoáteguiUbv MargaritaNessuna valutazione finora
- ABSUELTODocumento14 pagineABSUELTOJOSENessuna valutazione finora
- Regaxidil ProspectoDocumento5 pagineRegaxidil ProspectoAdela Martínez CamposNessuna valutazione finora
- Salud Mental: Cuidados Basicos Mente CuerpoDocumento2 pagineSalud Mental: Cuidados Basicos Mente Cuerporashell jimenezNessuna valutazione finora
- Edades FINAL Expandido Notas BookletDocumento226 pagineEdades FINAL Expandido Notas BookletLeiber TejenaNessuna valutazione finora
- El Pensamiento Pedagógico de Agustin YañezDocumento4 pagineEl Pensamiento Pedagógico de Agustin YañezJessica tIBBARNessuna valutazione finora
- Planeacion TangramDocumento3 paginePlaneacion TangramFloricelly Osorio Marfil60% (5)
- Acta #1 2017 - 2018 InicialDocumento2 pagineActa #1 2017 - 2018 InicialJosuéAltamiranoNessuna valutazione finora
- Novela Gotica y Romanticismo Oscuro PDFDocumento2 pagineNovela Gotica y Romanticismo Oscuro PDFHedera90100% (1)
- Tipos de Propagacion AsexualDocumento27 pagineTipos de Propagacion AsexualyuliNessuna valutazione finora
- Peritaje - Normas y Atribuciones Del PeritoDocumento127 paginePeritaje - Normas y Atribuciones Del PeritoSilvia Dionisio100% (1)