Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- PDFDocumento3 paginePDFPhan-128282Nessuna valutazione finora
- OHS Handbook Mar 03Documento111 pagineOHS Handbook Mar 03Phan-128282Nessuna valutazione finora
- JD Technician Operator BrewhouseDocumento3 pagineJD Technician Operator BrewhousePhan-128282Nessuna valutazione finora
- UTF 8ar syPT3024 1Documento64 pagineUTF 8ar syPT3024 1Phan-128282Nessuna valutazione finora
- 90259859 (2)Documento2 pagine90259859 (2)Phan-128282Nessuna valutazione finora
- Malts and Malting: Printed BookDocumento1 paginaMalts and Malting: Printed BookPhan-128282Nessuna valutazione finora
- A Real Time Conv PDFDocumento14 pagineA Real Time Conv PDFPhan-128282Nessuna valutazione finora
- A Real Time Conv PDFDocumento14 pagineA Real Time Conv PDFPhan-128282Nessuna valutazione finora
- Silva 01 Cement Summit Retarders3Documento1 paginaSilva 01 Cement Summit Retarders3Phan-128282Nessuna valutazione finora
- SystemC-Tutorial Paderborn PDFDocumento98 pagineSystemC-Tutorial Paderborn PDFPhan-128282Nessuna valutazione finora
- Convolutional Neural NetworkDocumento22 pagineConvolutional Neural NetworkPhan-128282100% (1)
- SystemC-Tutorial Paderborn PDFDocumento98 pagineSystemC-Tutorial Paderborn PDFPhan-128282Nessuna valutazione finora
- 2017 AppliedSurfaceScienceDocumento9 pagine2017 AppliedSurfaceSciencePhan-128282Nessuna valutazione finora
- Development of Modular Outdoor Furniture Product Using Lightweight Concrete For Public Parks in SurabayaDocumento8 pagineDevelopment of Modular Outdoor Furniture Product Using Lightweight Concrete For Public Parks in SurabayaPhan-128282Nessuna valutazione finora
- The Grinding-Aids Effect of Moisture, Triethanolamine (TEA) and Ethylene Glycol (EG) On Grinding Performance and Product Quality of CalciteDocumento8 pagineThe Grinding-Aids Effect of Moisture, Triethanolamine (TEA) and Ethylene Glycol (EG) On Grinding Performance and Product Quality of CalcitePhan-128282Nessuna valutazione finora
- Ban Ve Tong The Dan Quay TNDocumento1 paginaBan Ve Tong The Dan Quay TNPhan-128282Nessuna valutazione finora
- Silva 01 Cement-Summit-Retarders3 PDFDocumento16 pagineSilva 01 Cement-Summit-Retarders3 PDFb4rfNessuna valutazione finora
- BBBBDocumento5 pagineBBBBPhan-128282Nessuna valutazione finora
- Toolbox RácDocumento1 paginaToolbox RácPhan-128282Nessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- CodominanceDocumento23 pagineCodominanceKrizzel JuliahNessuna valutazione finora
- Health Status and Quality of Life of Geriatric Population in Old Age Homes and Living With Family in Chennai A Comparative StudyDocumento5 pagineHealth Status and Quality of Life of Geriatric Population in Old Age Homes and Living With Family in Chennai A Comparative StudyInternational Journal of Innovative Science and Research TechnologyNessuna valutazione finora
- Standard Operating Procedure For Somatic Variant Refinement of Sequencing Data With Paired Tumor and Normal SamplesDocumento10 pagineStandard Operating Procedure For Somatic Variant Refinement of Sequencing Data With Paired Tumor and Normal SamplesLaurie Stephens-KelleherNessuna valutazione finora
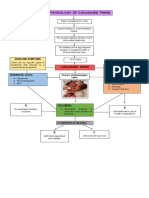
- Pathophysiology of Conjoined TwinsDocumento1 paginaPathophysiology of Conjoined TwinsPrincess Loreto AlegreNessuna valutazione finora
- 003 REVIEWER IN SCIENCE 7-SUgarDocumento3 pagine003 REVIEWER IN SCIENCE 7-SUgarrizza suarezNessuna valutazione finora
- Cichetti y Rogosh (2002) - Developmental Psychology PerspectiveDocumento16 pagineCichetti y Rogosh (2002) - Developmental Psychology PerspectiveAlejandro De la CruzNessuna valutazione finora
- Genevieve Balunis Resume 2017Documento3 pagineGenevieve Balunis Resume 2017Dakota BalunisNessuna valutazione finora
- Is Basic Technique Used For The Examination of Movement and For Initiating Movement Into A Program of Therapeutic InterventionDocumento9 pagineIs Basic Technique Used For The Examination of Movement and For Initiating Movement Into A Program of Therapeutic InterventionEric Carlos YumulNessuna valutazione finora
- CS 2000iDocumento88 pagineCS 2000iBintang MaulanaNessuna valutazione finora
- Syllabus - Biochemistry - 090216 PDFDocumento38 pagineSyllabus - Biochemistry - 090216 PDFchicklit subliminalNessuna valutazione finora
- Sample-Anatomy and Physiology The Unity of Form and Function 9th 9EDocumento30 pagineSample-Anatomy and Physiology The Unity of Form and Function 9th 9Erist0% (1)
- 6 The Main PeriodsDocumento5 pagine6 The Main PeriodsUtami MNessuna valutazione finora
- Ujmr 2020 - 4 - 24 - 30 PDFDocumento7 pagineUjmr 2020 - 4 - 24 - 30 PDFUMYU Journal of Microbiology Research (UJMR)Nessuna valutazione finora
- Long Range Sports Med 15Documento6 pagineLong Range Sports Med 15api-361293242Nessuna valutazione finora
- Q1 Module 3 Science 9Documento32 pagineQ1 Module 3 Science 9リリスセレインNessuna valutazione finora
- Immobilization of Catalase Via Adsorption Into NatDocumento6 pagineImmobilization of Catalase Via Adsorption Into NatErwin Guillermo Valdizón WinterNessuna valutazione finora
- 10.3 The Nerve ImpulseDocumento20 pagine10.3 The Nerve ImpulseazwelljohnsonNessuna valutazione finora
- Agrias Claudina Intermedia: Two New Records of From Eastern ColombiaDocumento2 pagineAgrias Claudina Intermedia: Two New Records of From Eastern ColombiaJuan Carlos Flores PazNessuna valutazione finora
- BAM Chapter 29. CronobacterDocumento14 pagineBAM Chapter 29. CronobacterremyNessuna valutazione finora
- Cuso4 PDFDocumento6 pagineCuso4 PDFSabrina ShenaNessuna valutazione finora
- MotivationDocumento2 pagineMotivationPrachi Srivastava100% (1)
- Boger Boenninghausen Characteristics RepertoryDocumento12 pagineBoger Boenninghausen Characteristics RepertoryDrx Rustom AliNessuna valutazione finora
- 2019 Haplogroup HDocumento182 pagine2019 Haplogroup HKlaus MarklNessuna valutazione finora
- Silviculture: February 2018Documento69 pagineSilviculture: February 2018Tang YinNessuna valutazione finora
- Beckers World of The Cell 9th Edition Hardin Test Bank DownloadDocumento23 pagineBeckers World of The Cell 9th Edition Hardin Test Bank Downloaddaddockstudderyxeq100% (27)
- Designing For The Human SensesDocumento50 pagineDesigning For The Human SensesRahaf MachaalNessuna valutazione finora
- Frog DissertationDocumento171 pagineFrog Dissertationcreatingcreator100% (1)
- 2019 Introduction To PharmacologyDocumento46 pagine2019 Introduction To PharmacologyJohn ParajoNessuna valutazione finora
- WRT 1020 Cause and Effect Paper - SmokingDocumento10 pagineWRT 1020 Cause and Effect Paper - Smokingapi-248441390Nessuna valutazione finora
- Experiments in Microbiology Plant Pathology and Biotechnology PDFDocumento2 pagineExperiments in Microbiology Plant Pathology and Biotechnology PDFDenise50% (8)