Potrebbero piacerti anche
- 02a SEXUAL REPRODUCTION IN FLOWERING PLANTSDocumento35 pagine02a SEXUAL REPRODUCTION IN FLOWERING PLANTSRitik Kumar NayakNessuna valutazione finora
- Transport in PlantsDocumento65 pagineTransport in Plantswienna1987Nessuna valutazione finora
- 12 Reproduction Biology Notes IGCSE PDFDocumento51 pagine12 Reproduction Biology Notes IGCSE PDFUmair Khan Marwat100% (1)
- EcologyDocumento32 pagineEcologyzaino4567123Nessuna valutazione finora
- Lecture notes-XII-Sexual Reproduction in Flowering PlantsDocumento11 pagineLecture notes-XII-Sexual Reproduction in Flowering Plantsaditya100% (3)
- Transport in PlantsDocumento6 pagineTransport in PlantsFred H HalderNessuna valutazione finora
- Quiz 1 J23 (1) - MergedDocumento16 pagineQuiz 1 J23 (1) - MergedmauoshaNessuna valutazione finora
- CELL STRUCTURE AND FUNCTIONS RecapDocumento17 pagineCELL STRUCTURE AND FUNCTIONS RecapSonakshi Chavan100% (1)
- Nutrition PlantDocumento3 pagineNutrition PlantKUOKNessuna valutazione finora
- Transport in Plants (Vascular Bundle)Documento6 pagineTransport in Plants (Vascular Bundle)Ahmed Kaleem Khan NiaziNessuna valutazione finora
- Homework 7 J23.Documento15 pagineHomework 7 J23.mauoshaNessuna valutazione finora
- Pollination in PlantsDocumento214 paginePollination in PlantsAbraham SekyiNessuna valutazione finora
- Cell Organelles WorksheetDocumento8 pagineCell Organelles WorksheetJohn OsborneNessuna valutazione finora
- IGCSE Answers Chapters 01 05Documento7 pagineIGCSE Answers Chapters 01 05María Eugenia MolteniNessuna valutazione finora
- Biology 1 - Characteristics of Living OrganismsDocumento79 pagineBiology 1 - Characteristics of Living OrganismsSaket GudimellaNessuna valutazione finora
- Cell-The Unit of LifeDocumento15 pagineCell-The Unit of LifeSiddhivinayak TidakeNessuna valutazione finora
- Mitosis and Cell CyclesDocumento12 pagineMitosis and Cell Cyclessg note0% (3)
- Animal Kingdom Notes: Vertebrates vs InvertebratesDocumento5 pagineAnimal Kingdom Notes: Vertebrates vs InvertebratesJacquiline Santiago100% (1)
- Biology Paper 2 (Question) LDocumento15 pagineBiology Paper 2 (Question) Lhasimahazit75% (8)
- Al Rowad International School, Riyadh: Biology IGCSE CambridgeDocumento154 pagineAl Rowad International School, Riyadh: Biology IGCSE CambridgeNicayne RamnarineNessuna valutazione finora
- Leaf Structure Function GuideDocumento1 paginaLeaf Structure Function GuideJasmine VS032689100% (1)
- Reproduction in Organisms: Asexual Reproduction and Life SpanDocumento11 pagineReproduction in Organisms: Asexual Reproduction and Life SpanRiya MondalNessuna valutazione finora
- Biology Chapter 4 Test ReviewDocumento7 pagineBiology Chapter 4 Test ReviewCK. GohNessuna valutazione finora
- Notes: - Reproduction in Organisms, For Class 12Documento18 pagineNotes: - Reproduction in Organisms, For Class 12Subho BhattacharyaNessuna valutazione finora
- Transportation in PlantsDocumento5 pagineTransportation in PlantsMitali PatelNessuna valutazione finora
- A2 4 3 1 Introduction To PhotosynthesisDocumento9 pagineA2 4 3 1 Introduction To PhotosynthesisDebbieJonesNessuna valutazione finora
- BiologyDocumento39 pagineBiologyAnna MontiniNessuna valutazione finora
- Cell Organelles WorksheetDocumento8 pagineCell Organelles Worksheet53777730% (2)
- Name: Date: Class: Instruction: For Each of The Questions 1 To 30, Choose The Correct Answer For Each Question From The Four Options A, B, C and DDocumento4 pagineName: Date: Class: Instruction: For Each of The Questions 1 To 30, Choose The Correct Answer For Each Question From The Four Options A, B, C and Dke2100% (1)
- Marking Schemes For Topic 1,2 Stafford PapersDocumento57 pagineMarking Schemes For Topic 1,2 Stafford PapersSuha AbdullahNessuna valutazione finora
- Chapter 11 Review PacketDocumento17 pagineChapter 11 Review PacketLalisampt.20% (1)
- Causes of Variation in SpeciesDocumento35 pagineCauses of Variation in SpeciesPatrick AbidraNessuna valutazione finora
- Heredity and VariationDocumento25 pagineHeredity and VariationDre ParkerNessuna valutazione finora
- Biology DefinitionsDocumento6 pagineBiology DefinitionsSixtine MorinNessuna valutazione finora
- Inheritance Exam QDocumento29 pagineInheritance Exam Qapi-422428700100% (1)
- Classification StudysheetDocumento36 pagineClassification Studysheetwafa eliasNessuna valutazione finora
- 9aa Environmental Variation VLE V3Documento32 pagine9aa Environmental Variation VLE V3syztqNessuna valutazione finora
- All About FlowersDocumento43 pagineAll About FlowersjeffinjoffiNessuna valutazione finora
- Variety of Living OrganismsDocumento4 pagineVariety of Living Organismsapi-255623302Nessuna valutazione finora
- MCQs of Plant TransportationDocumento5 pagineMCQs of Plant Transportationprofessor zulfi100% (2)
- Transportation of Food and Minerals in PlantsDocumento3 pagineTransportation of Food and Minerals in PlantsRonnith NandyNessuna valutazione finora
- Chapter 8.dynamic EcosystemDocumento45 pagineChapter 8.dynamic EcosystemNur Bahiyah Binti Abdul Wahab IPGKTINessuna valutazione finora
- IGCSE - Bio - Lesson Plan 13 - Reproduction in PlantsDocumento3 pagineIGCSE - Bio - Lesson Plan 13 - Reproduction in PlantsHisokagenNessuna valutazione finora
- Evaluation Sheet: Good Average Needs ImprovementDocumento15 pagineEvaluation Sheet: Good Average Needs ImprovementMalak ShokryNessuna valutazione finora
- Paper 2 November 1999Documento5 paginePaper 2 November 1999MSHNessuna valutazione finora
- Homework 5 J23Documento5 pagineHomework 5 J23mauoshaNessuna valutazione finora
- Chapter 3 Heredity and VariationDocumento14 pagineChapter 3 Heredity and Variationuncleya07100% (1)
- Microorganisms: Friends and FoeDocumento9 pagineMicroorganisms: Friends and FoeNIKHILPATNINessuna valutazione finora
- Human Reproduction Question BankDocumento23 pagineHuman Reproduction Question Bankkaustubhchoudhary2005Nessuna valutazione finora
- Chapter 15 Hormones & Endocrine Glands - Lecture NotesDocumento5 pagineChapter 15 Hormones & Endocrine Glands - Lecture Notesapi-3728508100% (3)
- Classification of Vertebrate and Invertebrate AnimalsDocumento33 pagineClassification of Vertebrate and Invertebrate AnimalsJanice Tañedo PancipaneNessuna valutazione finora
- 5 Kingdom ClassificationDocumento36 pagine5 Kingdom ClassificationMichelle Arienza100% (1)
- Biology Module Chap 8Documento37 pagineBiology Module Chap 8Faida Hamid100% (4)
- Transport in Plants NotesDocumento14 pagineTransport in Plants NotesUsama KhanNessuna valutazione finora
- O Level Biology Practice Questions And Answers Transport In PlantsDa EverandO Level Biology Practice Questions And Answers Transport In PlantsNessuna valutazione finora
- Reproduction in Lower and Higher PlantsDocumento89 pagineReproduction in Lower and Higher Plantsmohdyasir6643Nessuna valutazione finora
- Class 12 Subject: Biology Chapter-2: FlowerDocumento30 pagineClass 12 Subject: Biology Chapter-2: FlowerSufiyanNessuna valutazione finora
- Morphology of Flowering PlantsDocumento24 pagineMorphology of Flowering PlantsNalla Raghuram ChowdaryNessuna valutazione finora
- Bantham Hooker Classification System FinalDocumento1 paginaBantham Hooker Classification System FinalSumit KumarNessuna valutazione finora
- Division Spermatophyta 2012Documento68 pagineDivision Spermatophyta 2012tomatasiin10Nessuna valutazione finora
- Gymno AngioDocumento40 pagineGymno AngioBrainy headNessuna valutazione finora
- Lecture 5 Flower MorphologyDocumento39 pagineLecture 5 Flower MorphologyNur SabrinaNessuna valutazione finora
- Sexual Reproduction in Flowering Plants MCQs IDocumento12 pagineSexual Reproduction in Flowering Plants MCQs IMahendhiran MariappanNessuna valutazione finora
- LTDS2.2 Sexual Reproduction in Flowering PlantsDocumento8 pagineLTDS2.2 Sexual Reproduction in Flowering PlantsShane FernandezNessuna valutazione finora
- Reproduction in Plants Powerpoint SlidesDocumento55 pagineReproduction in Plants Powerpoint SlidesTay Kai xin (Ahs)Nessuna valutazione finora
- Chapter 3a - Reproduction in Plants - 3.1-3.4Documento18 pagineChapter 3a - Reproduction in Plants - 3.1-3.4Dorothy NgNessuna valutazione finora
- MLP HLP Class 7 Reproduction in PlantsDocumento5 pagineMLP HLP Class 7 Reproduction in Plantsbose baraniNessuna valutazione finora
- Sexual IncompatibilityDocumento8 pagineSexual IncompatibilitynipanshuNessuna valutazione finora
- Angiosperms and Their LifecycleDocumento48 pagineAngiosperms and Their LifecycleMnqobi ZuluNessuna valutazione finora
- Oxalis Del PERUDocumento290 pagineOxalis Del PERUpanchofrancoNessuna valutazione finora
- Flowering Plants: Life-Cycle of A Flowering PlantDocumento5 pagineFlowering Plants: Life-Cycle of A Flowering Plantraunak sarafNessuna valutazione finora
- Detailed Lesson Plan IN Science V: Prepared By: Checked byDocumento6 pagineDetailed Lesson Plan IN Science V: Prepared By: Checked byJosefina Trazona100% (12)
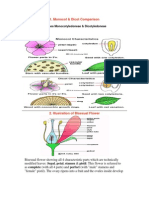
- Monocot & Dicot Comparison: Classes Monocotyledoneae & DicotyledoneaeDocumento4 pagineMonocot & Dicot Comparison: Classes Monocotyledoneae & DicotyledoneaeSyan LeeNessuna valutazione finora
- Sexual Reproduction in Flowering PlantsDocumento4 pagineSexual Reproduction in Flowering PlantsHarpreetNessuna valutazione finora
- DR - Ali Objective For BioDocumento345 pagineDR - Ali Objective For BioKashish Yadav100% (1)
- Chapter 12-Reproduction in PlantsDocumento4 pagineChapter 12-Reproduction in PlantsrajeshkumarkashyapNessuna valutazione finora
- Harvest Mice Love The Smell of Pollen and Often Fall Asleep Inside FlowersDocumento1 paginaHarvest Mice Love The Smell of Pollen and Often Fall Asleep Inside FlowersAndrew Richard ThompsonNessuna valutazione finora
- Plant Parts and Their FunctionsDocumento8 paginePlant Parts and Their FunctionsReanna Mae A. BaganiNessuna valutazione finora
- Kartveluri Memkvidreoba I Lomtadze, PutkaradzeDocumento4 pagineKartveluri Memkvidreoba I Lomtadze, PutkaradzeKARTVELIAN HERITAGE INessuna valutazione finora
- 12th Sci: ST. John's ClassesDocumento2 pagine12th Sci: ST. John's ClassesSHRI DEVKINANDAN JI BHAJANNessuna valutazione finora
- Angiosperms PPT RevisedDocumento55 pagineAngiosperms PPT RevisedJohn Philip Neri Besedillas100% (1)
- Perpetuation of Life PlantsDocumento28 paginePerpetuation of Life PlantsRosemarie EstoresNessuna valutazione finora
- Sexual Reproduction in Flowering PlantsDocumento66 pagineSexual Reproduction in Flowering Plantsshobitgup100% (1)
- How Weeds Spread Their SeedsDocumento3 pagineHow Weeds Spread Their SeedsCristina AguinaldoNessuna valutazione finora
- Q2 - Third Summative TestDocumento3 pagineQ2 - Third Summative TestMilagros ReyesNessuna valutazione finora
- Foraging BehaviorofhoneybeesDocumento6 pagineForaging BehaviorofhoneybeesWaqar HassanNessuna valutazione finora
- Anthurium Breeding PPT by S Y ChandrashekarDocumento30 pagineAnthurium Breeding PPT by S Y ChandrashekarDr-Chandrashekar S Y100% (20)