Potrebbero piacerti anche
- Bioquimica CUESTIONARIODocumento3 pagineBioquimica CUESTIONARIOSalvadorCerinza88% (8)
- 3 RESUMEN - Fosforilación OxidativaDocumento9 pagine3 RESUMEN - Fosforilación OxidativaMauricio DelgadilloNessuna valutazione finora
- DiastasaDocumento4 pagineDiastasaWALTER ADOLFO TACUCHE RIOSNessuna valutazione finora
- Vía de Las Pentosas FosfatoDocumento18 pagineVía de Las Pentosas FosfatoAndré HerreraNessuna valutazione finora
- Mapa Conceptual de Fosforilacion OxidativaDocumento2 pagineMapa Conceptual de Fosforilacion OxidativaGABRIELA ISABEL PACHERRES SALDA�A50% (2)
- Tipos de TraducciónDocumento3 pagineTipos de TraducciónLourdes Perdomo100% (1)
- Fase 3, CuestionarioDocumento34 pagineFase 3, CuestionarioSilvia Bustamante0% (1)
- Fosforilación OxidativaDocumento4 pagineFosforilación OxidativaDaleska C. GalarzaNessuna valutazione finora
- Cadena Transporte de Electrones y Fosforilacion OxidativaDocumento51 pagineCadena Transporte de Electrones y Fosforilacion OxidativaLuisNessuna valutazione finora
- Fosforilacion OxidativaDocumento31 pagineFosforilacion Oxidativadanieles93Nessuna valutazione finora
- Tema 7 - Fosforilación OxidativaDocumento13 pagineTema 7 - Fosforilación OxidativaLauraNessuna valutazione finora
- Fosforilacion OxidativaDocumento19 pagineFosforilacion OxidativaImanol De la SotaNessuna valutazione finora
- Transporte Electrónico y Fosforilación OxidativaDocumento30 pagineTransporte Electrónico y Fosforilación OxidativaPaola GVNessuna valutazione finora
- Cadena Electronic A y Fosforilacion OxidativaDocumento63 pagineCadena Electronic A y Fosforilacion OxidativaNoe Brito MorelNessuna valutazione finora
- GlucólisisDocumento26 pagineGlucólisislautarok100% (2)
- Tema 13 Ruta Pentosa-FosfatoDocumento6 pagineTema 13 Ruta Pentosa-FosfatoIsismar VillamizarNessuna valutazione finora
- Fosforilacion OxidativaDocumento5 pagineFosforilacion Oxidativaaoihana18100% (1)
- Cadena de Trasporte y Fosforilacion KKDocumento32 pagineCadena de Trasporte y Fosforilacion KKUriel AlvaradoNessuna valutazione finora
- Via de Las Pentosas Fosfato y Nadph (Harvey Unidad 13)Documento8 pagineVia de Las Pentosas Fosfato y Nadph (Harvey Unidad 13)IlderAlvaradoNessuna valutazione finora
- Ciclo de La Pentosa FosfatoDocumento7 pagineCiclo de La Pentosa FosfatoeiderNessuna valutazione finora
- Via de La Pentosa-FosfatoDocumento18 pagineVia de La Pentosa-FosfatoMarieta MOreira PincayNessuna valutazione finora
- La Ruta de Las Pentosas FosfatoDocumento47 pagineLa Ruta de Las Pentosas FosfatoSergio Emilio Fllres FuentesNessuna valutazione finora
- Sindrome de HunterDocumento18 pagineSindrome de HunterFernando Callata CardenasNessuna valutazione finora
- Fosforilacion OxidativaDocumento11 pagineFosforilacion OxidativaJose Alberto Ariza PasoNessuna valutazione finora
- Cadena Respiratoria Exposicion FinalDocumento28 pagineCadena Respiratoria Exposicion Finalesneider alexander perilla vergaraNessuna valutazione finora
- Vía de Las Pentosas FosfatoDocumento8 pagineVía de Las Pentosas Fosfatobrenda reynosoNessuna valutazione finora
- Ciclo de KrebsDocumento15 pagineCiclo de KrebsDaniel Saenz DominguezNessuna valutazione finora
- GlucogenólisisDocumento3 pagineGlucogenólisisJosué Delgado Navas100% (1)
- Ciclo de La GlucolisisDocumento13 pagineCiclo de La GlucolisisCriss AdrianNessuna valutazione finora
- T7. Cadena Respiratoria-CLAVDocumento53 pagineT7. Cadena Respiratoria-CLAVPaolo Gianfranco Luna Victoria GutierrezNessuna valutazione finora
- GLUCONEOGENESISDocumento32 pagineGLUCONEOGENESISzatanna66100% (2)
- Biosíntesis de Acidos GrasosDocumento2 pagineBiosíntesis de Acidos Grasosbryer92Nessuna valutazione finora
- Vía de Las Pentosas Fosfato y El CicloDocumento19 pagineVía de Las Pentosas Fosfato y El CicloGuadalupe Del Rocio Guerrero100% (1)
- GLUCÓLISISDocumento15 pagineGLUCÓLISISDylan EstebanNessuna valutazione finora
- Bioquimica Via de Pentosas FosfatoDocumento21 pagineBioquimica Via de Pentosas FosfatoLucero LópezNessuna valutazione finora
- TEMA 46 Biosintesis de Ácidos GrasosDocumento6 pagineTEMA 46 Biosintesis de Ácidos GrasosAkimita100% (3)
- Metabolismo de LipidosDocumento49 pagineMetabolismo de LipidosSheyla Espino RamosNessuna valutazione finora
- La FotosìntesisDocumento37 pagineLa FotosìntesiskarlaNessuna valutazione finora
- Tema13 Catabolismo AminoacidosDocumento36 pagineTema13 Catabolismo AminoacidosÁngela Díaz ChamorroNessuna valutazione finora
- Vía de Las Pentosas FosfatoDocumento19 pagineVía de Las Pentosas FosfatoAndré HerreraNessuna valutazione finora
- Rutas MetabolicasDocumento30 pagineRutas Metabolicasamilcar torresNessuna valutazione finora
- GlucolisisDocumento15 pagineGlucolisisalejo_1994244644100% (1)
- Genetica BacterianaDocumento75 pagineGenetica Bacterianaannys18Nessuna valutazione finora
- Ciclo de KrebsDocumento9 pagineCiclo de KrebsMariela Barrantes CorderoNessuna valutazione finora
- GlucogenolisisDocumento47 pagineGlucogenolisisDannerPlay25 20Nessuna valutazione finora
- Biología I Tema 3.2.5.3 Cadena RespiratoriaDocumento30 pagineBiología I Tema 3.2.5.3 Cadena RespiratoriaTita Oz100% (1)
- BIOQUIMICA Ciclo de KrebsDocumento74 pagineBIOQUIMICA Ciclo de KrebsleonelNessuna valutazione finora
- Fosforilación OxidativaDocumento25 pagineFosforilación OxidativaEmilio Jeanpaul Mitacc GutierrezNessuna valutazione finora
- MicosisDocumento9 pagineMicosisJimy Isaac S-romeroNessuna valutazione finora
- Biosíntesis de Carbohidratos - BIOQUIMICA II U2Documento13 pagineBiosíntesis de Carbohidratos - BIOQUIMICA II U2Bernardo Morales0% (1)
- Destinos Del PiruvatoDocumento29 pagineDestinos Del PiruvatoAdonai EsparzaNessuna valutazione finora
- Bioquimica Cadena RespiratoriaDocumento23 pagineBioquimica Cadena RespiratoriaMaríaCarolinaDiazgranadosGarcíaNessuna valutazione finora
- Tema 10 Bioquimica IIDocumento4 pagineTema 10 Bioquimica IIYolanda Chaves GuerreroNessuna valutazione finora
- Metabolismo de Los Seres VivosDocumento19 pagineMetabolismo de Los Seres VivosRoberto199524Nessuna valutazione finora
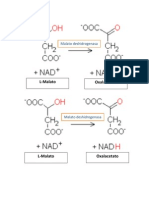
- La Enzima Malato DeshidrogenasaDocumento2 pagineLa Enzima Malato DeshidrogenasaKaren Torres JuárezNessuna valutazione finora
- Metabolismo de Las ProteínasDocumento40 pagineMetabolismo de Las ProteínasFrank Sánchez Estela100% (1)
- Cadena Respiratoria y Fosforilacion OxidativaDocumento21 pagineCadena Respiratoria y Fosforilacion OxidativaViridiana GonzalezNessuna valutazione finora
- Clase 6 Metabolismo de Ácidos NucleicosDocumento18 pagineClase 6 Metabolismo de Ácidos NucleicosAguss FernandezNessuna valutazione finora
- Oxidación de Ácidos Grasos.Documento23 pagineOxidación de Ácidos Grasos.José May100% (2)
- Resumen MicrosDocumento3 pagineResumen MicrosChristian Huerta100% (1)
- Cadena Transportadora de ElectronesDocumento4 pagineCadena Transportadora de ElectronesAndres CarreroNessuna valutazione finora
- Cadena Respiratoria y FosforilaciónDocumento6 pagineCadena Respiratoria y FosforilaciónStacy Horna AnguloNessuna valutazione finora
- Expo BiologíaDocumento4 pagineExpo BiologíalecthoraNessuna valutazione finora
- Edades QuirurgicasDocumento14 pagineEdades QuirurgicasLourdes PerdomoNessuna valutazione finora
- Neumonia PediatriaDocumento20 pagineNeumonia PediatriaLourdes PerdomoNessuna valutazione finora
- Suturas EliDocumento34 pagineSuturas EliLourdes PerdomoNessuna valutazione finora
- Hormona Del Crecimiento o Somatotropina (GH)Documento8 pagineHormona Del Crecimiento o Somatotropina (GH)Lourdes PerdomoNessuna valutazione finora
- Abd Agudo Quirurgico BqtoDocumento88 pagineAbd Agudo Quirurgico BqtoLourdes PerdomoNessuna valutazione finora
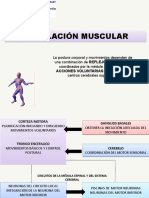
- Regulación Muscular1Documento19 pagineRegulación Muscular1Lourdes PerdomoNessuna valutazione finora
- Enfermedades Del Colágeno (SUBRAYADO)Documento13 pagineEnfermedades Del Colágeno (SUBRAYADO)Lourdes PerdomoNessuna valutazione finora
- Prosecucion de Estudios Año Escolar 2020-2021Documento10 pagineProsecucion de Estudios Año Escolar 2020-2021Lourdes PerdomoNessuna valutazione finora
- Objetivo 1 de InformaticaDocumento6 pagineObjetivo 1 de InformaticaLourdes PerdomoNessuna valutazione finora
- Hormona Del Crecimiento o Somatotropina (GH)Documento8 pagineHormona Del Crecimiento o Somatotropina (GH)Lourdes PerdomoNessuna valutazione finora
- Carácter Gobierno: Guerra Federal 1858 y 1863Documento28 pagineCarácter Gobierno: Guerra Federal 1858 y 1863Lourdes PerdomoNessuna valutazione finora
- Funcionamiento de Los MúsculosDocumento2 pagineFuncionamiento de Los MúsculosLourdes PerdomoNessuna valutazione finora
- Valencias FisicasDocumento5 pagineValencias FisicasLourdes PerdomoNessuna valutazione finora
- Circulación MayorDocumento8 pagineCirculación MayorLourdes PerdomoNessuna valutazione finora
- Disponibilidad para La Inclusion EscolarDocumento5 pagineDisponibilidad para La Inclusion EscolarLourdes PerdomoNessuna valutazione finora
- Centro RespiratorioDocumento3 pagineCentro RespiratorioLourdes PerdomoNessuna valutazione finora
- Glosario de Terminos DeportivoagilidadDocumento14 pagineGlosario de Terminos DeportivoagilidadLourdes PerdomoNessuna valutazione finora
- Valencias FisicasDocumento5 pagineValencias FisicasLourdes PerdomoNessuna valutazione finora
- Respiración Celular (Guia)Documento33 pagineRespiración Celular (Guia)Lourdes Perdomo100% (1)
- Clase Practica 27 Kenya Gil SDocumento9 pagineClase Practica 27 Kenya Gil SKenya SantacruzNessuna valutazione finora
- Sistemas Energéticos en El EjercicioDocumento65 pagineSistemas Energéticos en El EjercicioEdicson CASTANEDA POLANCONessuna valutazione finora
- Fosforilacion OxidativaDocumento11 pagineFosforilacion OxidativaJose Alberto Ariza PasoNessuna valutazione finora
- Muerte DulceDocumento29 pagineMuerte Dulcejacobo clavijoNessuna valutazione finora
- Guía #5 Metabolismo Aeróbico CQU310Documento6 pagineGuía #5 Metabolismo Aeróbico CQU310Jeane Nicole Poggi ArancibiaNessuna valutazione finora
- Tema 2 BioenergeticaDocumento17 pagineTema 2 BioenergeticaCristhian Macalupú Arismendiz100% (1)
- Microbiologia Parcial 2Documento33 pagineMicrobiologia Parcial 2mikichiko2349Nessuna valutazione finora
- Cianuro en El OrganismoDocumento4 pagineCianuro en El OrganismoBautista GioNessuna valutazione finora
- Respiracion CelularDocumento46 pagineRespiracion CelularMiguel Fierro0% (1)
- P1 Morfofisiología IDocumento13 pagineP1 Morfofisiología IIsbelis Flores100% (3)
- Bioquimica de La Obesidad Des2016Documento25 pagineBioquimica de La Obesidad Des2016Roxana Magaly Chavez Pillaca100% (1)
- Respiración CelularDocumento16 pagineRespiración CelularDionar Ruiz altamiranoNessuna valutazione finora
- Principios Energéticos y Producción de EnergíaDocumento13 paginePrincipios Energéticos y Producción de EnergíaDosty Veliz100% (2)
- Cadena RespiratoriaDocumento25 pagineCadena RespiratoriaKruzkaya MachicadoNessuna valutazione finora
- La SimbiosisDocumento8 pagineLa Simbiosisdaht19Nessuna valutazione finora
- Examen Resuelto BiolI B4Documento18 pagineExamen Resuelto BiolI B4Mari Marisol Rodriguez50% (2)
- CAPITULO 5 BIOQUIMICA II (1) EliDocumento32 pagineCAPITULO 5 BIOQUIMICA II (1) EliAlberth MuñozNessuna valutazione finora
- T-10 Teoria QuimiosmóticaDocumento30 pagineT-10 Teoria QuimiosmóticaUlises Benjamin Narro Altamirano0% (1)
- AE007 BioquimicaDocumento23 pagineAE007 BioquimicaJuanca PradoNessuna valutazione finora
- The ERK and JNK Pathways in The Regulation of Metabolic - En.es - En.esDocumento18 pagineThe ERK and JNK Pathways in The Regulation of Metabolic - En.es - En.esAngi Carolina Valdelamar MonteroNessuna valutazione finora
- Seminario BioquímicaDocumento34 pagineSeminario BioquímicaStacy Horna AnguloNessuna valutazione finora
- CITOLOGIADocumento27 pagineCITOLOGIAdeieuNessuna valutazione finora
- 5 Tema VI Guía HC Krebs y Cadena Respiratoria 2019Documento4 pagine5 Tema VI Guía HC Krebs y Cadena Respiratoria 2019valeNessuna valutazione finora
- Clase 2012 8 9 Metabolismo de CHO PDFDocumento128 pagineClase 2012 8 9 Metabolismo de CHO PDFAngie Paola Padua CalleNessuna valutazione finora
- Ejercicio 2 - Tarea 3 - Grupo Colaborativo - 44Documento22 pagineEjercicio 2 - Tarea 3 - Grupo Colaborativo - 44Julieth BarahonaNessuna valutazione finora
- CASO CLÍNICO 5 Intoxicacion X CODocumento16 pagineCASO CLÍNICO 5 Intoxicacion X COClaudiaGabrielaMirandaConsuegraNessuna valutazione finora
- Bombeo de Protones en Membrana Celular de Levaduras y Sus Procesos InhibitoriosDocumento7 pagineBombeo de Protones en Membrana Celular de Levaduras y Sus Procesos InhibitoriosGabriela CoyNessuna valutazione finora
- SEGUNDO PERIODO Biologia 1Documento5 pagineSEGUNDO PERIODO Biologia 1Zahhy BonnieNessuna valutazione finora