Potrebbero piacerti anche
- JMV 25888-2Documento8 pagineJMV 25888-2Cynthia Torres-GonzálezNessuna valutazione finora
- Attachment Based Psychotherapy Helping Patients Develop Adaptive Capacities PDFDocumento281 pagineAttachment Based Psychotherapy Helping Patients Develop Adaptive Capacities PDFCynthia Torres-GonzálezNessuna valutazione finora
- Conte. Emotion Knowledge, ToMDocumento11 pagineConte. Emotion Knowledge, ToMCynthia Torres-GonzálezNessuna valutazione finora
- Caillaud Et Al-2019-Human Brain MappingDocumento11 pagineCaillaud Et Al-2019-Human Brain MappingCynthia Torres-GonzálezNessuna valutazione finora
- Fnhum 13 00271Documento7 pagineFnhum 13 00271Cynthia Torres-GonzálezNessuna valutazione finora
- Boccadoro. Defining The Neural Correlates of Spontaneous Theory of MindDocumento9 pagineBoccadoro. Defining The Neural Correlates of Spontaneous Theory of MindCynthia Torres-GonzálezNessuna valutazione finora
- Cannon 1927Documento20 pagineCannon 1927Cynthia Torres-GonzálezNessuna valutazione finora
- JCM 08 01086Documento9 pagineJCM 08 01086Cynthia Torres-GonzálezNessuna valutazione finora
- Green Et Al-2019-Autism ResearchDocumento14 pagineGreen Et Al-2019-Autism ResearchCynthia Torres-GonzálezNessuna valutazione finora
- Caillaud Et Al-2019-Human Brain MappingDocumento11 pagineCaillaud Et Al-2019-Human Brain MappingCynthia Torres-GonzálezNessuna valutazione finora
- Clearfield. SES Affects Infant Cognitive FlexibilityDocumento7 pagineClearfield. SES Affects Infant Cognitive FlexibilityCynthia Torres-GonzálezNessuna valutazione finora
- Cole. The Closing ToMDocumento16 pagineCole. The Closing ToMCynthia Torres-GonzálezNessuna valutazione finora
- The "Reading The Mind in The Eyes" Test Revised Version - A Study With Normal Adults, and Adults With Asperger Syndrome or High-Functioning Autis PDFDocumento11 pagineThe "Reading The Mind in The Eyes" Test Revised Version - A Study With Normal Adults, and Adults With Asperger Syndrome or High-Functioning Autis PDFIrfan FauziNessuna valutazione finora
- Caillaud Et Al-2019-Human Brain MappingDocumento11 pagineCaillaud Et Al-2019-Human Brain MappingCynthia Torres-GonzálezNessuna valutazione finora
- Appropriateness and Limitations of Factor Analysis Methods Utilised in Psychology and Kinesiology-Part 2Documento13 pagineAppropriateness and Limitations of Factor Analysis Methods Utilised in Psychology and Kinesiology-Part 2Cynthia Torres-GonzálezNessuna valutazione finora
- Bon. Growing Up With Asperger SyndromeDocumento8 pagineBon. Growing Up With Asperger SyndromeCynthia Torres-GonzálezNessuna valutazione finora
- Theory of Mind Training Improves Social Skills in Children with AutismDocumento10 pagineTheory of Mind Training Improves Social Skills in Children with AutismCynthia Torres-GonzálezNessuna valutazione finora
- Bowman. Contintuity in The Neural System Supporting Childrens ToM DevelopmentDocumento10 pagineBowman. Contintuity in The Neural System Supporting Childrens ToM DevelopmentCynthia Torres-GonzálezNessuna valutazione finora
- LibroDocumento251 pagineLibroCynthia Torres-GonzálezNessuna valutazione finora
- G Model Study Examines Age-Related Changes in Cognitive and Affective Components of Theory of MindDocumento11 pagineG Model Study Examines Age-Related Changes in Cognitive and Affective Components of Theory of MindCynthia Torres-GonzálezNessuna valutazione finora
- Classifying Emotion: A Developmental Account: Alexandra Zinck Albert NewenDocumento25 pagineClassifying Emotion: A Developmental Account: Alexandra Zinck Albert NewenCynthia Torres-GonzálezNessuna valutazione finora
- Impact of Age On Emotional Intelligence and Its Components: Deeksha SharmaDocumento8 pagineImpact of Age On Emotional Intelligence and Its Components: Deeksha SharmaCynthia Torres-GonzálezNessuna valutazione finora
- Altered Functional Connectivity of The Posterior Cingulate Cortez in Type 2 DiabetesDocumento9 pagineAltered Functional Connectivity of The Posterior Cingulate Cortez in Type 2 DiabetesCynthia Torres-GonzálezNessuna valutazione finora
- ArticuloDocumento21 pagineArticuloCynthia Torres-GonzálezNessuna valutazione finora
- This Is Chapter 1 From The Book, Cognition in Emotion, by Tone Roald, Published by Brill in 2007Documento14 pagineThis Is Chapter 1 From The Book, Cognition in Emotion, by Tone Roald, Published by Brill in 2007Cynthia Torres-GonzálezNessuna valutazione finora
- Creativity As Predictor of Mathematical Abilities in Fourth Graders in Addition To Number Sense and Working MemoryDocumento24 pagineCreativity As Predictor of Mathematical Abilities in Fourth Graders in Addition To Number Sense and Working MemoryCynthia Torres-GonzálezNessuna valutazione finora
- Buddhism's Influence on Psychology and NeuroscienceDocumento16 pagineBuddhism's Influence on Psychology and NeuroscienceCynthia Torres-GonzálezNessuna valutazione finora
- PNAS 2017 LeDoux 1619316114Documento10 paginePNAS 2017 LeDoux 1619316114Luis PastorNessuna valutazione finora
- Affective Consciousness: Core Emotional Feelings in Animals and HumansDocumento51 pagineAffective Consciousness: Core Emotional Feelings in Animals and HumansCynthia Torres-GonzálezNessuna valutazione finora
- The Basic Emotional Circuits of Mammalia PDFDocumento14 pagineThe Basic Emotional Circuits of Mammalia PDFCynthia Torres-GonzálezNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Otology Fellowships 2019Documento5 pagineOtology Fellowships 2019Sandra SandrinaNessuna valutazione finora
- Organs of Speech GuideDocumento14 pagineOrgans of Speech GuideJen N. de ChavezNessuna valutazione finora
- Descending Tracts: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeDocumento39 pagineDescending Tracts: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeÑäd ÉèmNessuna valutazione finora
- Dysphagia After Stroke-An Overview PDFDocumento18 pagineDysphagia After Stroke-An Overview PDFDhea SafitriNessuna valutazione finora
- MRI & CT of Facial Bones PDFDocumento53 pagineMRI & CT of Facial Bones PDFdr_340802071Nessuna valutazione finora
- Termin Root Prefix SuffixDocumento3 pagineTermin Root Prefix SuffixAkun CanvaNessuna valutazione finora
- He International Frontal Sinus Anatomy Classification (IFAC) and Classification of The Extent of Endoscopic Frontal Sinus Surgery (EFSSDocumento20 pagineHe International Frontal Sinus Anatomy Classification (IFAC) and Classification of The Extent of Endoscopic Frontal Sinus Surgery (EFSSanitaabreu123Nessuna valutazione finora
- The Middle Ear TransformerDocumento13 pagineThe Middle Ear TransformerDiego RozulNessuna valutazione finora
- LEC M3.3 BlockADocumento8 pagineLEC M3.3 BlockASieg100% (1)
- 05 - CSF + Ventricles - DR Najeeb VideosDocumento26 pagine05 - CSF + Ventricles - DR Najeeb Videoshiba jasimNessuna valutazione finora
- Introduction to Prosthodontics and Complete DenturesDocumento86 pagineIntroduction to Prosthodontics and Complete DenturesKamal NidaNessuna valutazione finora
- Neck Pain Chronic Sinusitis and Eustachian Tube Dysfunction - Caring Medical FloridaDocumento2 pagineNeck Pain Chronic Sinusitis and Eustachian Tube Dysfunction - Caring Medical FloridaTulisanNessuna valutazione finora
- Hypothalamus-Pituitary-Thyroid Axis: PRESENTERS: DR: Nyangaresi Justine DR: Mitchelle Facilitator: DRDocumento31 pagineHypothalamus-Pituitary-Thyroid Axis: PRESENTERS: DR: Nyangaresi Justine DR: Mitchelle Facilitator: DRJustine NyangaresiNessuna valutazione finora
- ENT-Laryngeal ParalysisDocumento26 pagineENT-Laryngeal ParalysisRahul Kumar DiwakarNessuna valutazione finora
- Kalpa in The PhysiologyDocumento58 pagineKalpa in The PhysiologysaidevoNessuna valutazione finora
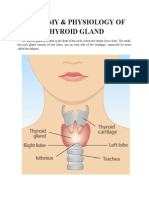
- Anatomy & Physiology of Thyroid GlandDocumento5 pagineAnatomy & Physiology of Thyroid GlandJohnpaul DelapenaNessuna valutazione finora
- Full Download Test Bank For Sectional Anatomy For Imaging Professionals 3rd Edition Kelley PDF Full ChapterDocumento36 pagineFull Download Test Bank For Sectional Anatomy For Imaging Professionals 3rd Edition Kelley PDF Full Chapterflotageepigee.bp50100% (23)
- Zundorf 2015 Dual Pathway Auditory SystemDocumento10 pagineZundorf 2015 Dual Pathway Auditory SystemRicardo Jose De LeonNessuna valutazione finora
- The Cricoid CartilageDocumento1 paginaThe Cricoid CartilageStasinopoulou Maria ElisavetNessuna valutazione finora
- Digestive and Respiratory Organs DefinitionDocumento4 pagineDigestive and Respiratory Organs DefinitionSidne E. AgorNessuna valutazione finora
- Inhibitory Control Over Action and MemoryDocumento11 pagineInhibitory Control Over Action and MemoryMaryam TaqaviNessuna valutazione finora
- L1) Oral Cavity, Palate and TongueDocumento39 pagineL1) Oral Cavity, Palate and Tongue3amroooni.a.aNessuna valutazione finora
- Thyroid Disease OutlineDocumento3 pagineThyroid Disease OutlinePaige BarniniNessuna valutazione finora
- Trans Savi Oto Lec 01 Head and Neck History and PE 1st SemesterDocumento12 pagineTrans Savi Oto Lec 01 Head and Neck History and PE 1st SemesterJoherNessuna valutazione finora
- Thyroid Colloid Nodular GoiterDocumento23 pagineThyroid Colloid Nodular Goitervahn025Nessuna valutazione finora
- MandibleDocumento120 pagineMandibleArnab SantraNessuna valutazione finora
- Nanda 2016Documento15 pagineNanda 2016Novia AyuNessuna valutazione finora
- Apraxia, Agnosia and Disconnection SyndromesDocumento65 pagineApraxia, Agnosia and Disconnection SyndromesDhawal Narang100% (1)
- Retrosigmoid Approach for Resecting Petroclival MeningiomasDocumento7 pagineRetrosigmoid Approach for Resecting Petroclival MeningiomasCAMILO ARMANDO BENAVIDES BURBANONessuna valutazione finora
- Dokumen - Pub - Sensory Integration and Learning Disorders Hardcovernbsped 0874243033 9780874243031 - Compressed 11Documento12 pagineDokumen - Pub - Sensory Integration and Learning Disorders Hardcovernbsped 0874243033 9780874243031 - Compressed 11Pablo Rodríguez HernándezNessuna valutazione finora