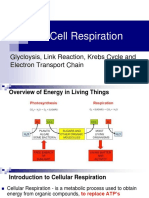
Potrebbero piacerti anche
- Respiration in PlantsDocumento6 pagineRespiration in PlantsjanakiramannadesanNessuna valutazione finora
- BioenergeticsDocumento29 pagineBioenergeticskizzaymenteraNessuna valutazione finora
- Cellular Respiration PDFDocumento10 pagineCellular Respiration PDFAnonymous HXLczq3Nessuna valutazione finora
- 1.4 Respiration-The Source of EnergyDocumento14 pagine1.4 Respiration-The Source of EnergysamiktshyagiriNessuna valutazione finora
- 2-Energy and RespirationDocumento34 pagine2-Energy and RespirationLisa DentonNessuna valutazione finora
- Chapter 14 - RESPIRATION IN PLANTSDocumento10 pagineChapter 14 - RESPIRATION IN PLANTSprernatiwary508Nessuna valutazione finora
- Biology NotesDocumento214 pagineBiology NotesbloomNessuna valutazione finora
- Regulation of glycolysis to meet energy demandsDocumento7 pagineRegulation of glycolysis to meet energy demandsFatmata Haja KamaraNessuna valutazione finora
- Cellular Respiration - The Energy-Producing Process in Living CellsDocumento21 pagineCellular Respiration - The Energy-Producing Process in Living CellsAbirami SankarNessuna valutazione finora
- Metabolism BKSDocumento26 pagineMetabolism BKSAjmain ShakibNessuna valutazione finora
- Cellular Respiration ExplanationDocumento2 pagineCellular Respiration Explanationvestermail50% (2)
- Cellular Metabolism ExplainedDocumento14 pagineCellular Metabolism ExplainedAndre NesbethNessuna valutazione finora
- Respiration GuideDocumento30 pagineRespiration GuideSamuel HarvNessuna valutazione finora
- Powerpoint To Accompany Concepts in Biology: Twelfth Edition Enger - Ross - BaileyDocumento24 paginePowerpoint To Accompany Concepts in Biology: Twelfth Edition Enger - Ross - BaileyAndika HidayatullahNessuna valutazione finora
- Earth and Life Science SHS 16.3 How Organisms Obtain and Utilize EnergyDocumento23 pagineEarth and Life Science SHS 16.3 How Organisms Obtain and Utilize EnergyPamela MalihanNessuna valutazione finora
- Chapter 7_ Introduction to Cellular RespirationDocumento14 pagineChapter 7_ Introduction to Cellular Respirationbabylen bahalaNessuna valutazione finora
- How Cells Release Energy Part 1Documento14 pagineHow Cells Release Energy Part 1Jiro SarmientoNessuna valutazione finora
- Biology 2nd QRTR Exam - OutlineDocumento5 pagineBiology 2nd QRTR Exam - OutlineLouie Joice MartinezNessuna valutazione finora
- A2 Biology Notes Cellular RespirationDocumento20 pagineA2 Biology Notes Cellular RespirationArnel100% (1)
- Las Q4 WK3.1Documento8 pagineLas Q4 WK3.1Perlyn Del Pilar OduyaNessuna valutazione finora
- Carbohydrate MetabolismDocumento30 pagineCarbohydrate MetabolismWycliff MuchaiNessuna valutazione finora
- Plant Respiration ExplainedDocumento6 paginePlant Respiration ExplainedAadil ShakulNessuna valutazione finora
- RespirationDocumento100 pagineRespirationColin RiddellNessuna valutazione finora
- Reviewer-In-Gen Bio 1 - Q2Documento5 pagineReviewer-In-Gen Bio 1 - Q2alamagelyzaNessuna valutazione finora
- CH 6 Cellular RespirationDocumento28 pagineCH 6 Cellular RespirationANessuna valutazione finora
- Course Title: Course Code:: First Capital University of Bangladesh, ChuadangaDocumento17 pagineCourse Title: Course Code:: First Capital University of Bangladesh, Chuadangamanik sarkarNessuna valutazione finora
- CMB Merge 2Documento137 pagineCMB Merge 2Jr RamosNessuna valutazione finora
- CBSE Class-11 Biology Revision - Respiration in PlantsDocumento4 pagineCBSE Class-11 Biology Revision - Respiration in PlantsSHRAVYA K REDDYNessuna valutazione finora
- BIOENERGETICSDocumento3 pagineBIOENERGETICSJaemsNessuna valutazione finora
- Lecture 7 Respiration in Plants_glycoysis_fermentation_oxidative Phosphorylation PptDocumento31 pagineLecture 7 Respiration in Plants_glycoysis_fermentation_oxidative Phosphorylation Pptmichael328wanjiruNessuna valutazione finora
- Chemistru Final ProjectDocumento26 pagineChemistru Final ProjectSanskar MohanNessuna valutazione finora
- Bio 102 Exam Concept List Cell Organelles Respiration PhotosynthesisDocumento3 pagineBio 102 Exam Concept List Cell Organelles Respiration PhotosynthesisLanae BraswellNessuna valutazione finora
- Cellular RespirationDocumento10 pagineCellular RespirationXavier LecarosNessuna valutazione finora
- Glycolysis and the Citric Acid CycleDocumento6 pagineGlycolysis and the Citric Acid CycleNhư NguyễnNessuna valutazione finora
- Introduction To Cellular RespirationDocumento5 pagineIntroduction To Cellular RespirationCarlton GrantNessuna valutazione finora
- Understanding the Metabolic Processes of CarbohydratesDocumento20 pagineUnderstanding the Metabolic Processes of CarbohydratesAHKI ANTONETTE ABELNessuna valutazione finora
- Chapter 14Documento14 pagineChapter 14Yash Singh 11th BNessuna valutazione finora
- Cell RespirationDocumento29 pagineCell RespirationJohn OsborneNessuna valutazione finora
- CarbohydratesDocumento19 pagineCarbohydratesNaina YadavvNessuna valutazione finora
- Cellular - Respiration Lecture KoDocumento60 pagineCellular - Respiration Lecture KoEvangelene Esquillo SanaNessuna valutazione finora
- Cellular Respiration NotesDocumento10 pagineCellular Respiration Notescsujithan100% (1)
- Carbohydrate Metabolism 2Documento20 pagineCarbohydrate Metabolism 2kanikichoniNessuna valutazione finora
- Hsslive-xi-botany-RESPIRATION IN PLANTS-8Documento7 pagineHsslive-xi-botany-RESPIRATION IN PLANTS-8diyakozhimalaNessuna valutazione finora
- Campbell Biology Concepts and Connections 9th Edition Taylor Solutions ManualDocumento11 pagineCampbell Biology Concepts and Connections 9th Edition Taylor Solutions Manualkevinrodriguezcnytfpaskx100% (22)
- Ebook Campbell Biology Concepts and Connections 9Th Edition Taylor Solutions Manual Full Chapter PDFDocumento32 pagineEbook Campbell Biology Concepts and Connections 9Th Edition Taylor Solutions Manual Full Chapter PDFtristandoyledtnpkozbgj100% (11)
- Cellular Respiration (MAEd Assign)Documento4 pagineCellular Respiration (MAEd Assign)jajajaredredNessuna valutazione finora
- HMS 4601 NUTRITION TO FUEL SPORTS PERFORMANCE - Case 1Documento23 pagineHMS 4601 NUTRITION TO FUEL SPORTS PERFORMANCE - Case 1stijn.berkmortel1Nessuna valutazione finora
- Science 9 q1w7Documento9 pagineScience 9 q1w7seanfelix07102009Nessuna valutazione finora
- Place Homework Into The Basket. 2. Pick Up Powerpoint GuideDocumento52 paginePlace Homework Into The Basket. 2. Pick Up Powerpoint GuideADR GARDIOLANessuna valutazione finora
- Revision Handout 1 For Grade 12Documento10 pagineRevision Handout 1 For Grade 12babaNessuna valutazione finora
- Cell RespirationDocumento29 pagineCell RespirationJohn OsborneNessuna valutazione finora
- NEET Respiration Notes - Glycolysis, Fermentation, Krebs CycleDocumento6 pagineNEET Respiration Notes - Glycolysis, Fermentation, Krebs CycleThahseen AfzalNessuna valutazione finora
- NEET Respiration Notes: Glycolysis, Fermentation, Kreb's CycleDocumento6 pagineNEET Respiration Notes: Glycolysis, Fermentation, Kreb's CycleThahseen AfzalNessuna valutazione finora
- Jinnah University Women Aerobic AnaerobicDocumento13 pagineJinnah University Women Aerobic AnaerobicDEVIL GAMINGNessuna valutazione finora
- General Biology 1: Quarter 2 - Module 13 To 15: Energy TransformationDocumento8 pagineGeneral Biology 1: Quarter 2 - Module 13 To 15: Energy Transformationkent ignacioNessuna valutazione finora
- Universidad Autónoma de Nuevo León School of Chemistry.: Microbiology Part 2: Metabolism and GrowthDocumento12 pagineUniversidad Autónoma de Nuevo León School of Chemistry.: Microbiology Part 2: Metabolism and GrowthCarlos Cerón LinaresNessuna valutazione finora
- Lecture 13. Introduction To Metabolism: - Catabolism - Degradation - Anabolism (Aka Biosynthesis) - EnergyDocumento20 pagineLecture 13. Introduction To Metabolism: - Catabolism - Degradation - Anabolism (Aka Biosynthesis) - EnergyAnonymous Bh9NrU77Nessuna valutazione finora
- Bio Lesson 21 RespiDocumento48 pagineBio Lesson 21 RespiChlzy ArdienteNessuna valutazione finora
- Cellular Respiration - Aerobic and AnaerobicDocumento5 pagineCellular Respiration - Aerobic and AnaerobicJeaña Despi DaguismolNessuna valutazione finora
- Multiple page document scanned by CamScannerDocumento29 pagineMultiple page document scanned by CamScannereddula ganeshNessuna valutazione finora
- Complex VariablesDocumento29 pagineComplex Variableseddula ganeshNessuna valutazione finora
- Partial Differencial OperatiosDocumento38 paginePartial Differencial Operatioseddula ganeshNessuna valutazione finora
- MT Notes Part 1 Nageswar Sir NotesDocumento8 pagineMT Notes Part 1 Nageswar Sir Noteseddula ganeshNessuna valutazione finora
- MT Part 4 Class Notes Nageswar SirDocumento66 pagineMT Part 4 Class Notes Nageswar Sireddula ganeshNessuna valutazione finora
- Complex ContinuationDocumento21 pagineComplex Continuationeddula ganeshNessuna valutazione finora
- Fourier Series Part 2Documento20 pagineFourier Series Part 2eddula ganeshNessuna valutazione finora
- MT Part 2 Nageswar Sir NotesDocumento42 pagineMT Part 2 Nageswar Sir Noteseddula ganeshNessuna valutazione finora
- MT Mid To EndDocumento74 pagineMT Mid To Endeddula ganeshNessuna valutazione finora
- MT Part 4 Class Notes Nageswar SirDocumento66 pagineMT Part 4 Class Notes Nageswar Sireddula ganeshNessuna valutazione finora
- MT Previous PaperDocumento2 pagineMT Previous Papereddula ganeshNessuna valutazione finora
- MT Part 2 Nageswar Sir NotesDocumento42 pagineMT Part 2 Nageswar Sir Noteseddula ganeshNessuna valutazione finora
- " Physical Metallurgy Laboratory": Lab Report OnDocumento1 pagina" Physical Metallurgy Laboratory": Lab Report Oneddula ganeshNessuna valutazione finora
- MT Notes Part 3 Nageswar Sir NotesDocumento38 pagineMT Notes Part 3 Nageswar Sir Noteseddula ganeshNessuna valutazione finora
- MT Notes Part 3 Nageswar Sir NotesDocumento38 pagineMT Notes Part 3 Nageswar Sir Noteseddula ganeshNessuna valutazione finora
- MT Previous PaperDocumento2 pagineMT Previous Papereddula ganeshNessuna valutazione finora
- BEE Lab Manual-ModifiedDocumento30 pagineBEE Lab Manual-Modifiededdula ganeshNessuna valutazione finora
- Free Course and Programming Guide PDFDocumento50 pagineFree Course and Programming Guide PDFSTOKYNessuna valutazione finora
- KineticsDocumento14 pagineKineticseddula ganeshNessuna valutazione finora
- Galvanic Cells AssignmentDocumento14 pagineGalvanic Cells Assignmenteddula ganeshNessuna valutazione finora
- Unit - 3 Pretreatment of Hot Metal PDFDocumento34 pagineUnit - 3 Pretreatment of Hot Metal PDFeddula ganeshNessuna valutazione finora
- MT Notes Part 1 Nageswar Sir NotesDocumento8 pagineMT Notes Part 1 Nageswar Sir Noteseddula ganeshNessuna valutazione finora
- Maths Till KrishDocumento13 pagineMaths Till Krisheddula ganeshNessuna valutazione finora
- " Physical Metallurgy Laboratory": Lab Report OnDocumento1 pagina" Physical Metallurgy Laboratory": Lab Report Oneddula ganeshNessuna valutazione finora
- Chemical Kinetics TopicDocumento14 pagineChemical Kinetics Topiceddula ganeshNessuna valutazione finora
- Taylor and Laurent Series for Complex FunctionsDocumento106 pagineTaylor and Laurent Series for Complex Functionseddula ganeshNessuna valutazione finora
- To ReviseDocumento1 paginaTo Reviseeddula ganeshNessuna valutazione finora
- New Things Learnt in ContestsDocumento1 paginaNew Things Learnt in Contestseddula ganeshNessuna valutazione finora
- Maths Till KrishnastamiDocumento9 pagineMaths Till Krishnastamieddula ganeshNessuna valutazione finora
- EE235 BEE Title Page and ListDocumento2 pagineEE235 BEE Title Page and Listeddula ganeshNessuna valutazione finora
- E-Mix - EASI Screed PDFDocumento3 pagineE-Mix - EASI Screed PDFngNessuna valutazione finora
- Epofine 337 - Finehard 3918Documento4 pagineEpofine 337 - Finehard 3918Karishma PrabhuNessuna valutazione finora
- Oh 2002Documento6 pagineOh 2002tehtnicaNessuna valutazione finora
- Sindh University 2014Documento9 pagineSindh University 2014Raees BachaNessuna valutazione finora
- Histopathology questions (AHC diplomaDocumento11 pagineHistopathology questions (AHC diplomaSAMMY89% (9)
- Chanchal SirDocumento38 pagineChanchal SirTouhid HossainNessuna valutazione finora
- Lab Report Steam DistillationDocumento6 pagineLab Report Steam DistillationBerdigylychNessuna valutazione finora
- Physico-Chemical Study of Seed Oil of Prunus Armeniaca L. Grown in Garhwal Region (India) and Its Comparison With Some Conventional Food OilsDocumento6 paginePhysico-Chemical Study of Seed Oil of Prunus Armeniaca L. Grown in Garhwal Region (India) and Its Comparison With Some Conventional Food OilsTIMESTAMP MASTERNessuna valutazione finora
- SOP - Portable Gas ChromatographyDocumento19 pagineSOP - Portable Gas ChromatographyKishanNessuna valutazione finora
- ISO 9963 1 1994 en PreviewDocumento3 pagineISO 9963 1 1994 en PreviewDeangelis DamasNessuna valutazione finora
- Pesticides List 2Documento564 paginePesticides List 2Ruth LeeNessuna valutazione finora
- EM 1110-1-4012 PrecipitationDocumento101 pagineEM 1110-1-4012 PrecipitationKenindra PranidyaNessuna valutazione finora
- Astm F880MDocumento5 pagineAstm F880MJUAN DAVID GOMEZ PATIÑONessuna valutazione finora
- 5 - GEST 10 362 Edition 4 - Corrosion Behaviour of Carbon Steel in Wet and Dry ChlorineDocumento41 pagine5 - GEST 10 362 Edition 4 - Corrosion Behaviour of Carbon Steel in Wet and Dry ChlorineAdemilson JuniorNessuna valutazione finora
- 013-DSV 10 - DSV 1 - DSV2Documento13 pagine013-DSV 10 - DSV 1 - DSV2john jimenezNessuna valutazione finora
- E572-13 - Standard Test Method For Analysis of Stainless and Alloy Steels by Wavelength Dispersive X-Ray Fluorescence Spectrometry.Documento11 pagineE572-13 - Standard Test Method For Analysis of Stainless and Alloy Steels by Wavelength Dispersive X-Ray Fluorescence Spectrometry.Royce BergNessuna valutazione finora
- GST English PCDocumento64 pagineGST English PCsharkrameshNessuna valutazione finora
- Bab6 FunctionDocumento25 pagineBab6 Functionbellaputri125fNessuna valutazione finora
- MBH2007Documento34 pagineMBH2007Azi GallantNessuna valutazione finora
- BCS /SS - CRM No. 405/2 Low Alloy Steel: B A S LDocumento2 pagineBCS /SS - CRM No. 405/2 Low Alloy Steel: B A S LPeterson SudlabNessuna valutazione finora
- Photosynthesis 7thDocumento33 paginePhotosynthesis 7thSatyam SinghNessuna valutazione finora
- TQP and TQSMDocumento23 pagineTQP and TQSMSalman QadirNessuna valutazione finora
- Antoine LavoisierDocumento9 pagineAntoine LavoisierLeo HuangNessuna valutazione finora
- METE 256 Lecture 1 - AssayingDocumento21 pagineMETE 256 Lecture 1 - AssayingGideon MbabilaNessuna valutazione finora
- PNP - List of PNP Controlled ChemicalsDocumento1 paginaPNP - List of PNP Controlled ChemicalsjajaNessuna valutazione finora
- 2ND Quarter Grade 9 ExaminationDocumento4 pagine2ND Quarter Grade 9 ExaminationSher Sherwin100% (1)
- Standard procedures for disinfectants against COVID-19Documento27 pagineStandard procedures for disinfectants against COVID-19andualemNessuna valutazione finora
- Struktur Materi GenetikDocumento73 pagineStruktur Materi GenetikFikry SyahrizalNessuna valutazione finora
- Specification For Polyaluminium Chloride - Lpac - Used in Potable Water Supply PDFDocumento6 pagineSpecification For Polyaluminium Chloride - Lpac - Used in Potable Water Supply PDFMOHD JIDINessuna valutazione finora
- FRP Unlimited: Connecting Professionals To The Value of Fibre Reinforced PlasticsDocumento5 pagineFRP Unlimited: Connecting Professionals To The Value of Fibre Reinforced Plasticsargentino_ar01Nessuna valutazione finora