Potrebbero piacerti anche
- Determinação Da Taxa Mínima de Armadura de Flexão em Vigas Com Seção RetangularDocumento9 pagineDeterminação Da Taxa Mínima de Armadura de Flexão em Vigas Com Seção RetangularPatrícia MouraNessuna valutazione finora
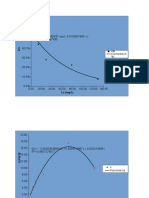
- Co (MG/L) : A% Exponential (A %)Documento35 pagineCo (MG/L) : A% Exponential (A %)Οδυσσεας ΚοψιδαςNessuna valutazione finora
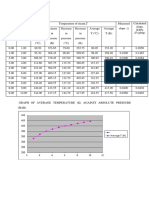
- Quality Check KI K2 K3 Sample Molwt Charge (MG/L) (Mmol/l) (MG/L) (Mmol/l (MG/L) (Mmol/l)Documento6 pagineQuality Check KI K2 K3 Sample Molwt Charge (MG/L) (Mmol/l) (MG/L) (Mmol/l (MG/L) (Mmol/l)CarolineMwitaMoseregaNessuna valutazione finora
- Weibull FinalDocumento10 pagineWeibull FinalJorge Paul Ortiz NuñezNessuna valutazione finora
- Co (MG/L) : A% Exponential (A %)Documento35 pagineCo (MG/L) : A% Exponential (A %)Οδυσσεας ΚοψιδαςNessuna valutazione finora
- Fogler 8.16Documento3 pagineFogler 8.16ivanaNessuna valutazione finora
- ZUCSDocumento9 pagineZUCSAndres GarciaNessuna valutazione finora
- Co (MG/L) : A% Exponential (A %)Documento35 pagineCo (MG/L) : A% Exponential (A %)Οδυσσεας ΚοψιδαςNessuna valutazione finora
- ECV 513 AssignmentDocumento15 pagineECV 513 AssignmentCarolineMwitaMoseregaNessuna valutazione finora
- Highway Alignment-R3Documento17 pagineHighway Alignment-R3TarunNessuna valutazione finora
- 08 - Chapter 3Documento39 pagine08 - Chapter 3adil rezaNessuna valutazione finora
- Calculation of Cut Petroleum FractionDocumento6 pagineCalculation of Cut Petroleum FractionEsra BelhajNessuna valutazione finora
- Steam Tables: F FG G o 3Documento3 pagineSteam Tables: F FG G o 3fery150283Nessuna valutazione finora
- Tugas 2 Thermo - Kelompok 1 - ArgonDocumento14 pagineTugas 2 Thermo - Kelompok 1 - ArgonkevinNessuna valutazione finora
- Odysseas Kopsidas - 8Documento35 pagineOdysseas Kopsidas - 8Οδυσσεας ΚοψιδαςNessuna valutazione finora
- Evol. Glucosa: LN (X) Conc Glucosa (G/L)Documento19 pagineEvol. Glucosa: LN (X) Conc Glucosa (G/L)Carlos Andres Rios BarraNessuna valutazione finora
- Co (MG/L) : A% Exponential (A %)Documento35 pagineCo (MG/L) : A% Exponential (A %)Οδυσσεας ΚοψιδαςNessuna valutazione finora
- 123dok Pengaruh+Total+Solid+Dan+Total+Alkali+Aktif+Pada+Black+Liqour+ (Lindi+Hitam) +Terhadap+Kualitas+Pulp+YDocumento51 pagine123dok Pengaruh+Total+Solid+Dan+Total+Alkali+Aktif+Pada+Black+Liqour+ (Lindi+Hitam) +Terhadap+Kualitas+Pulp+Ypathun tambaNessuna valutazione finora
- Odysseas Kopsidas - 12Documento35 pagineOdysseas Kopsidas - 12Οδυσσεας ΚοψιδαςNessuna valutazione finora
- Simple PendulumDocumento13 pagineSimple Pendulumthelmamusonda60Nessuna valutazione finora
- Module 1: Laboratory: Assignment - 1Documento6 pagineModule 1: Laboratory: Assignment - 1Siddharth PatelNessuna valutazione finora
- Libro 1Documento8 pagineLibro 1Oskr Wero NoriegaNessuna valutazione finora
- Practical Considerations For Conductivity and Total Dissolved Solids MeasurementDocumento5 paginePractical Considerations For Conductivity and Total Dissolved Solids MeasurementVarun KumarNessuna valutazione finora
- ChipsDocumento7 pagineChipsDannyNessuna valutazione finora
- Libro 1Documento3 pagineLibro 1Eva HerreraNessuna valutazione finora
- Steam Tables: F FG G o 3Documento2 pagineSteam Tables: F FG G o 3Sandi AslanNessuna valutazione finora
- v (RPM) v (RPS) ζ (pulg.onza) Pot exp (W) Nre NpoDocumento10 paginev (RPM) v (RPS) ζ (pulg.onza) Pot exp (W) Nre NpoYeferson Andy Alexis Chuchon GomezNessuna valutazione finora
- Turbulent Flow: Experimental Data and ResultsDocumento4 pagineTurbulent Flow: Experimental Data and ResultsSilviu ProNessuna valutazione finora
- BinnerDocumento2 pagineBinnerAsdy PutraNessuna valutazione finora
- BinnerDocumento2 pagineBinnerAsdy PutraNessuna valutazione finora
- Odysseas Kopsidas - 5Documento35 pagineOdysseas Kopsidas - 5Οδυσσεας ΚοψιδαςNessuna valutazione finora
- Steam TableDocumento5 pagineSteam TableAyush BhadauriaNessuna valutazione finora
- Hagedorn Brown CorrelationDocumento28 pagineHagedorn Brown Correlationberkah haniNessuna valutazione finora
- Experiment #1 Raynold Number: S/N Volume of Water Time Q (m3/s) V M/sDocumento7 pagineExperiment #1 Raynold Number: S/N Volume of Water Time Q (m3/s) V M/sMir Masood ShahNessuna valutazione finora
- R Lab 1Documento11 pagineR Lab 1Ahoud AlhaimliNessuna valutazione finora
- Lab Report Experiment 1Documento12 pagineLab Report Experiment 1Mohd Sayfull33% (3)
- Ejercicio 2Documento10 pagineEjercicio 2Joel Elias Cruz CondoriNessuna valutazione finora
- Tabel Hasil Perhitungan (Nozzle) : Grafik P Terhadap QDocumento5 pagineTabel Hasil Perhitungan (Nozzle) : Grafik P Terhadap QRobby Ilma FermanaNessuna valutazione finora
- Viscosity of c5h802 - 28Documento1 paginaViscosity of c5h802 - 28CharlesNessuna valutazione finora
- 2Cu2O + O2 (G) 4cuo T Deltah Deltas Deltag K Log (K) K Kcal Cal/K KcalDocumento3 pagine2Cu2O + O2 (G) 4cuo T Deltah Deltas Deltag K Log (K) K Kcal Cal/K Kcaldiegoj2hotmail.comNessuna valutazione finora
- Diagrama de EllinghanDocumento3 pagineDiagrama de Ellinghandiegoj2hotmail.comNessuna valutazione finora
- Determination of The Conductance of Strong and Weak ElectrolyteDocumento10 pagineDetermination of The Conductance of Strong and Weak Electrolytetotorost100% (12)
- Odysseas Kopsidas - 9Documento35 pagineOdysseas Kopsidas - 9Οδυσσεας ΚοψιδαςNessuna valutazione finora
- Odysseas Kopsidas - 7Documento35 pagineOdysseas Kopsidas - 7Οδυσσεας ΚοψιδαςNessuna valutazione finora
- Thermodynamic Properties of Polyethylene: R. M. C. PetreeDocumento10 pagineThermodynamic Properties of Polyethylene: R. M. C. PetreeRitesh AgarwalNessuna valutazione finora
- Kurva Kalibrasi: Hasil Pengamatan Dan PerhitunganDocumento11 pagineKurva Kalibrasi: Hasil Pengamatan Dan PerhitunganVito Naufal PriyoNessuna valutazione finora
- F (X) 0.7698461538x + 0.1584553846 R 0.97804078: Qrot (m3/h)Documento1 paginaF (X) 0.7698461538x + 0.1584553846 R 0.97804078: Qrot (m3/h)marius8840Nessuna valutazione finora
- Aluminium Conductor Composite Core ACCCDocumento5 pagineAluminium Conductor Composite Core ACCCAmlanshankar Deb BarmaNessuna valutazione finora
- Result Marcet 2Documento4 pagineResult Marcet 2Ahmed Qutb AkmalNessuna valutazione finora
- Result Marcet 2Documento4 pagineResult Marcet 2Ahmed Qutb AkmalNessuna valutazione finora
- Assignm Net 4 SolDocumento14 pagineAssignm Net 4 SolRhedo.AbdNessuna valutazione finora
- ENGRDocumento36 pagineENGRjherryNessuna valutazione finora
- T°c X Y: Benceno - Tolueno A 1 AtmDocumento3 pagineT°c X Y: Benceno - Tolueno A 1 AtmMaxi Iglesias Scott KennedyNessuna valutazione finora
- Common Exhaust Gas C C CDocumento3 pagineCommon Exhaust Gas C C CKhristel PenoliarNessuna valutazione finora
- Equilibrio de Fases (Benceno/Metanol) 1. Utilización de Software (Chemcad) Por RaoultDocumento15 pagineEquilibrio de Fases (Benceno/Metanol) 1. Utilización de Software (Chemcad) Por RaoultrubenNessuna valutazione finora
- Equilibrio de Fases (Benceno/Metanol) 1. Utilización de Software (Chemcad) Por RaoultDocumento15 pagineEquilibrio de Fases (Benceno/Metanol) 1. Utilización de Software (Chemcad) Por RaoultrubenNessuna valutazione finora
- Tabel Perencanaan Kuva Sectional Area (Csa) : MasukanDocumento10 pagineTabel Perencanaan Kuva Sectional Area (Csa) : MasukanZarkasi ManshurNessuna valutazione finora
- (Eagan Press Handbook Series. - Practical Guides For The Food Industry) Francis, Frederick John - Colorants Handbook-Amer Assn of Cereal Chemists - Eagan Press (1999) PDFDocumento142 pagine(Eagan Press Handbook Series. - Practical Guides For The Food Industry) Francis, Frederick John - Colorants Handbook-Amer Assn of Cereal Chemists - Eagan Press (1999) PDFStefania DavidNessuna valutazione finora
- OtooDocumento11 pagineOtooStefania DavidNessuna valutazione finora
- (Chemistry Research and Applications) Fitzgerald, Adrienne - Emulsifiers - Properties, Functions, and Applications-Nova Science Publishers (2015) PDFDocumento116 pagine(Chemistry Research and Applications) Fitzgerald, Adrienne - Emulsifiers - Properties, Functions, and Applications-Nova Science Publishers (2015) PDFStefania DavidNessuna valutazione finora
- Pectina ApaDocumento11 paginePectina ApaStefania DavidNessuna valutazione finora
- Ginger Oleoresin Chemical Composition, Bioactivity and Application As Bio-PreservativesDocumento8 pagineGinger Oleoresin Chemical Composition, Bioactivity and Application As Bio-PreservativesStefania DavidNessuna valutazione finora
- PDFDocumento3 paginePDFStefania DavidNessuna valutazione finora
- Effects of PolarDocumento9 pagineEffects of PolarStefania DavidNessuna valutazione finora
- Eur 20831 enDocumento13 pagineEur 20831 enStefania DavidNessuna valutazione finora
- Roy Fuller (Auth.) - Probiotics - The Scientific Basis-Springer Netherlands (1992)Documento405 pagineRoy Fuller (Auth.) - Probiotics - The Scientific Basis-Springer Netherlands (1992)Stefania David100% (1)
- Food Research International: Cláudia Puerari, Karina Teixeira Magalhães, Rosane Freitas SchwanDocumento7 pagineFood Research International: Cláudia Puerari, Karina Teixeira Magalhães, Rosane Freitas SchwanStefania DavidNessuna valutazione finora
- Huang 2013Documento10 pagineHuang 2013Stefania DavidNessuna valutazione finora
- Animal Nutrition: Leon BroomDocumento4 pagineAnimal Nutrition: Leon BroomStefania DavidNessuna valutazione finora
- Technology of Production of Edible Flours and Protein Products From Soybeans..Documento14 pagineTechnology of Production of Edible Flours and Protein Products From Soybeans..Stefania DavidNessuna valutazione finora
- Manufacturing of Sulfuric Acid by Lead Chamber Process and Contact ProcessDocumento14 pagineManufacturing of Sulfuric Acid by Lead Chamber Process and Contact ProcessDian Eka FajriyantoNessuna valutazione finora
- Sulphur and Its CompoundsDocumento13 pagineSulphur and Its CompoundsElizabeth OwenNessuna valutazione finora
- Oxygen Concentrator: How They WorkDocumento4 pagineOxygen Concentrator: How They WorkALEXANDRE SANTOSNessuna valutazione finora
- Analisis Kualitas Udara DI Kawasan ParkiDocumento10 pagineAnalisis Kualitas Udara DI Kawasan ParkiTry Ayu LestariNessuna valutazione finora
- Nitrox Diver ManualDocumento61 pagineNitrox Diver ManualskinerNessuna valutazione finora
- Sap-2 Material Balance: Basis: Unit Hour of ProductionDocumento17 pagineSap-2 Material Balance: Basis: Unit Hour of ProductionNandha KumarNessuna valutazione finora
- IUPAC Provisional RecommendationsDocumento3 pagineIUPAC Provisional Recommendationsdamyan petrovNessuna valutazione finora
- My TestDocumento15 pagineMy TestkrisnuNessuna valutazione finora
- Drilling ExampleDocumento200 pagineDrilling ExampleLuis Gustavo Choque BordaNessuna valutazione finora
- HYPERBARICDocumento16 pagineHYPERBARICJihad Elias ChahlaNessuna valutazione finora
- Declaration Acc. REACHDocumento65 pagineDeclaration Acc. REACHJulioNessuna valutazione finora
- Group 16 Short Notes Dexterchem A852B9N5aEFjR1MPDocumento20 pagineGroup 16 Short Notes Dexterchem A852B9N5aEFjR1MPGully GamingNessuna valutazione finora
- Oxygen Therapy - Dr. Satish DeopujariDocumento46 pagineOxygen Therapy - Dr. Satish DeopujaricrplzNessuna valutazione finora
- Sulphuric AcidDocumento4 pagineSulphuric AcidKhai AzNessuna valutazione finora
- Perkins Engine SAEDocumento7 paginePerkins Engine SAEsyamsul ashariNessuna valutazione finora
- Centrometal PelTec Kotao Na Pelete Tehnicke UputeDocumento109 pagineCentrometal PelTec Kotao Na Pelete Tehnicke UputelagusterNessuna valutazione finora
- JIPO Catalogue CrucibleDocumento1 paginaJIPO Catalogue CrucibleSNP suvananNessuna valutazione finora
- Shacman Servic ManualDocumento251 pagineShacman Servic ManualAlvin Siew Wai100% (2)
- GAS Measurement/ Alarm Equipments: Toka Seiki Co.,LtdDocumento4 pagineGAS Measurement/ Alarm Equipments: Toka Seiki Co.,LtdpcatruongNessuna valutazione finora
- Manufacturing Sulphuric AcidDocumento1 paginaManufacturing Sulphuric AcidSatria HalimNessuna valutazione finora
- Prefixes and Suffixes Crossword: Name: - Date: - PeriodDocumento1 paginaPrefixes and Suffixes Crossword: Name: - Date: - PeriodAri SuhendraNessuna valutazione finora
- Group 16 Oxygen and SulphurDocumento33 pagineGroup 16 Oxygen and SulphurMaureen DireroNessuna valutazione finora
- A Study On Sulfuric AcidDocumento3 pagineA Study On Sulfuric AcidChing BanKai XiangNessuna valutazione finora
- Pidato Bahasa InggrisDocumento2 paginePidato Bahasa InggrisDewi EndahNessuna valutazione finora
- Claus Plants Prove FlexibleDocumento3 pagineClaus Plants Prove Flexiblebakhtiari_afNessuna valutazione finora
- Con, Vs So3 % - Density of Sulfuric Acid and Sulfur TrioxideDocumento9 pagineCon, Vs So3 % - Density of Sulfuric Acid and Sulfur TrioxideAnonymous JWI6rqtNessuna valutazione finora
- Sulfuric Acid and OleumDocumento9 pagineSulfuric Acid and Oleumpatrickstar155Nessuna valutazione finora
- KING 200 Oxygen SysytemDocumento25 pagineKING 200 Oxygen SysytemMiiguelNessuna valutazione finora
- Oath Hub Leh PKGDocumento3 pagineOath Hub Leh PKGPiyush AgrawalNessuna valutazione finora
- PAT KIMIA 12 GENAP 2023 (Jawaban)Documento37 paginePAT KIMIA 12 GENAP 2023 (Jawaban)elikNessuna valutazione finora