Potrebbero piacerti anche
- Buttocks Lift For Tight ThighsDocumento17 pagineButtocks Lift For Tight ThighsLink ZeldaNessuna valutazione finora
- 8 Staff - 201508270829016160Documento1 pagina8 Staff - 201508270829016160Link ZeldaNessuna valutazione finora
- BV RhizaDocumento27 pagineBV RhizaLink ZeldaNessuna valutazione finora
- Analysis of A Protocol For The Treatment of Buttocks FolliculitisDocumento5 pagineAnalysis of A Protocol For The Treatment of Buttocks FolliculitisLink ZeldaNessuna valutazione finora
- Qualifiers 19Documento60 pagineQualifiers 19Link ZeldaNessuna valutazione finora
- Commerzbank Aktiengesellschaft Final Terms: Dated 4 July 2016 Relating ToDocumento52 pagineCommerzbank Aktiengesellschaft Final Terms: Dated 4 July 2016 Relating ToLink ZeldaNessuna valutazione finora
- 2017 Godwin PHD CH 1Documento28 pagine2017 Godwin PHD CH 1Link ZeldaNessuna valutazione finora
- Cui Heuven ICPhS2011Documento4 pagineCui Heuven ICPhS2011Link ZeldaNessuna valutazione finora
- 2018 Geurds, Prehistory Southern Central AmericaDocumento20 pagine2018 Geurds, Prehistory Southern Central AmericaLink ZeldaNessuna valutazione finora
- Context-Free Grammars: A Grammar Is A Set of Rules For Putting Strings Together and So Corresponds To A LanguageDocumento20 pagineContext-Free Grammars: A Grammar Is A Set of Rules For Putting Strings Together and So Corresponds To A LanguageLink ZeldaNessuna valutazione finora
- Dudden Hill SopnDocumento1 paginaDudden Hill SopnLink ZeldaNessuna valutazione finora
- Vogelaar&Resing 2018Documento18 pagineVogelaar&Resing 2018Link ZeldaNessuna valutazione finora
- Scope of CoverageDocumento4 pagineScope of CoverageLink ZeldaNessuna valutazione finora
- Stad2019 Article TheRoleOfCognitiveFlexibilityIDocumento24 pagineStad2019 Article TheRoleOfCognitiveFlexibilityILink ZeldaNessuna valutazione finora
- Common Coupled Fixed Points For Two Pairs of W - Compatible Maps in Partial - Metric SpacesDocumento22 pagineCommon Coupled Fixed Points For Two Pairs of W - Compatible Maps in Partial - Metric SpacesLink ZeldaNessuna valutazione finora
- Time-Reverse Modelling of Acoustic Wave Propagation in Attenuating MediaDocumento12 pagineTime-Reverse Modelling of Acoustic Wave Propagation in Attenuating MediaLink ZeldaNessuna valutazione finora
- Process Assessment in Dynamic Testing Using Electronic TangiblesDocumento16 pagineProcess Assessment in Dynamic Testing Using Electronic TangiblesLink ZeldaNessuna valutazione finora
- Official Contest RulesDocumento3 pagineOfficial Contest RulesLink ZeldaNessuna valutazione finora
- KH25L12835F PDFDocumento101 pagineKH25L12835F PDFLink ZeldaNessuna valutazione finora
- VA Ext 5 2014 Eng PDFDocumento1 paginaVA Ext 5 2014 Eng PDFLink ZeldaNessuna valutazione finora
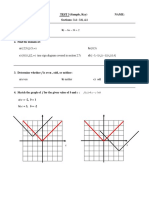
- MATH 153 TEST 3 (Sample, Key) Name: Sections: 3.4 - 3.8, 4.1Documento4 pagineMATH 153 TEST 3 (Sample, Key) Name: Sections: 3.4 - 3.8, 4.1Link ZeldaNessuna valutazione finora
- Wa0004Documento5 pagineWa0004Jms QuinteroNessuna valutazione finora
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5795)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Dharam Prakash SaranDocumento89 pagineDharam Prakash SarankeerthanaaNessuna valutazione finora
- Genetics 22-23Documento9 pagineGenetics 22-23hiNessuna valutazione finora
- Chapter 1 2 3Documento77 pagineChapter 1 2 3Ferissa MohammadNessuna valutazione finora
- 2.induction of Labour & Prolonged PregnancyDocumento20 pagine2.induction of Labour & Prolonged PregnancyMiswar Abdul HalimNessuna valutazione finora
- Lecture 3Documento39 pagineLecture 3Anne XxNessuna valutazione finora
- Raynaud's PhenomenonDocumento10 pagineRaynaud's PhenomenonRoberto López MataNessuna valutazione finora
- Anaphy Module 1Documento5 pagineAnaphy Module 110. Briol AlvinNessuna valutazione finora
- Red Flags For The MRCGPDocumento3 pagineRed Flags For The MRCGPUsm LeeNessuna valutazione finora
- Life ProcessesDocumento36 pagineLife Processes2erwr100% (1)
- 4.cardivascular System ExaminationDocumento60 pagine4.cardivascular System ExaminationElvisNessuna valutazione finora
- 26 Biology 2 - 16 - 08 Cellular RespirationDocumento48 pagine26 Biology 2 - 16 - 08 Cellular RespirationNiranjan BhuvanaratnamNessuna valutazione finora
- Pa Tho Physiology of GlomerulonephritisDocumento1 paginaPa Tho Physiology of GlomerulonephritisJhaziel BermejoNessuna valutazione finora
- Systemic Sclerosis SclerodermaDocumento40 pagineSystemic Sclerosis Sclerodermadrkorenofir100% (1)
- Braden Scale FormDocumento1 paginaBraden Scale FormJILL ANGELESNessuna valutazione finora
- Stages of SleepDocumento16 pagineStages of SleepMelanie100% (1)
- NSG Process-Chitra MamDocumento44 pagineNSG Process-Chitra MamJalajarani AridassNessuna valutazione finora
- Cardiac Arrest in The Operating Room (2012)Documento6 pagineCardiac Arrest in The Operating Room (2012)patricia.untoroNessuna valutazione finora
- Cranial Nerve Exam Handout 1Documento4 pagineCranial Nerve Exam Handout 1kingpin@mailinatorNessuna valutazione finora
- Kidney Anatomy and PhysiologyDocumento6 pagineKidney Anatomy and PhysiologyJohn Wilbert BirungNessuna valutazione finora
- WC500022748 2 PDFDocumento0 pagineWC500022748 2 PDFMostofa RubalNessuna valutazione finora
- Singing Technique and Anatomy Part 1Documento5 pagineSinging Technique and Anatomy Part 1musicguy joeNessuna valutazione finora
- BCA Protocol PDFDocumento7 pagineBCA Protocol PDFAlyssa236Nessuna valutazione finora
- Diabetes Mellitus IIDocumento20 pagineDiabetes Mellitus IIMa R Dy100% (1)
- BIO1 - Study Guide 4.4 - Cell Transport Mechanism II - Bulk or Vesicular TransportDocumento13 pagineBIO1 - Study Guide 4.4 - Cell Transport Mechanism II - Bulk or Vesicular TransportJustin JaranillaNessuna valutazione finora
- Biophysical ProfileDocumento26 pagineBiophysical ProfilePatricia Ysabelle Ledesma100% (1)
- 10 Current Research Articles Related To Our SystemDocumento11 pagine10 Current Research Articles Related To Our Systemhomework solutionNessuna valutazione finora
- Daftar List Ecatalog ABN 2022 With Pict Ok-2Documento5 pagineDaftar List Ecatalog ABN 2022 With Pict Ok-2SESILIA UTAMINessuna valutazione finora
- ArticlesDocumento4 pagineArticleschris7982Nessuna valutazione finora
- David Hartley S Psychobiological Associationism and The Legacy of AristotleDocumento18 pagineDavid Hartley S Psychobiological Associationism and The Legacy of AristotleA HollisNessuna valutazione finora
- Insomnia ABAUDocumento2 pagineInsomnia ABAURosa Pérez ParaparNessuna valutazione finora