Potrebbero piacerti anche
- Efectos de Las Heladas Sobre Los CultivosDocumento2 pagineEfectos de Las Heladas Sobre Los CultivosRuth Milagros Ruiz RiosNessuna valutazione finora
- Sindromes Secundarios - VihDocumento1 paginaSindromes Secundarios - VihRuth Milagros Ruiz RiosNessuna valutazione finora
- Caso 2Documento4 pagineCaso 2Ruth Milagros Ruiz RiosNessuna valutazione finora
- VERTEBRADOSDocumento3 pagineVERTEBRADOSRuth Milagros Ruiz RiosNessuna valutazione finora
- Objetivos y ConclDocumento2 pagineObjetivos y ConclRuth Milagros Ruiz RiosNessuna valutazione finora
- MONITORIZACIÓNDocumento3 pagineMONITORIZACIÓNRuth Milagros Ruiz RiosNessuna valutazione finora
- Taenia SoliumDocumento4 pagineTaenia SoliumRuth Milagros Ruiz RiosNessuna valutazione finora
- Heladas en Regiones Del PerúDocumento3 pagineHeladas en Regiones Del PerúRuth Milagros Ruiz RiosNessuna valutazione finora
- INTRODUCCIONSDocumento2 pagineINTRODUCCIONSRuth Milagros Ruiz RiosNessuna valutazione finora
- Historia Natural TBCDocumento5 pagineHistoria Natural TBCRuth Milagros Ruiz RiosNessuna valutazione finora
- ¿Cuáles Son Las Características de Patrón Restrictivo y Obstructivo en Una Espirometría?Documento5 pagine¿Cuáles Son Las Características de Patrón Restrictivo y Obstructivo en Una Espirometría?Ruth Milagros Ruiz RiosNessuna valutazione finora
- CONCLUSIONES - Preg 1 y 2Documento2 pagineCONCLUSIONES - Preg 1 y 2Ruth Milagros Ruiz RiosNessuna valutazione finora
- Clase Mammalia - Investigación TareaDocumento1 paginaClase Mammalia - Investigación TareaRuth Milagros Ruiz RiosNessuna valutazione finora
- Respuesta de Plantas Ante Bajas Temperaturas: Capítulo I Heladas Y FriajeDocumento4 pagineRespuesta de Plantas Ante Bajas Temperaturas: Capítulo I Heladas Y FriajeRuth Milagros Ruiz RiosNessuna valutazione finora
- Cuestionario 1,2,3 y ObjetivosDocumento3 pagineCuestionario 1,2,3 y ObjetivosRuth Milagros Ruiz RiosNessuna valutazione finora
- Problemas de Mutaciones GénicasDocumento3 pagineProblemas de Mutaciones GénicasRuth Milagros Ruiz RiosNessuna valutazione finora
- Biofisica ForoDocumento1 paginaBiofisica ForoRuth Milagros Ruiz RiosNessuna valutazione finora
- Pregunta 4 y 5 y ConclusionesDocumento2 paginePregunta 4 y 5 y ConclusionesRuth Milagros Ruiz RiosNessuna valutazione finora
- Tarea 13 Preg.05Documento2 pagineTarea 13 Preg.05Ruth Milagros Ruiz RiosNessuna valutazione finora
- OBJETIVOS, 9 y 10Documento2 pagineOBJETIVOS, 9 y 10Ruth Milagros Ruiz RiosNessuna valutazione finora
- Expo Punto 3Documento3 pagineExpo Punto 3Ruth Milagros Ruiz RiosNessuna valutazione finora
- Tarea 3 BoicaDocumento2 pagineTarea 3 BoicaRuth Milagros Ruiz RiosNessuna valutazione finora
- TAREA 5 BiocaDocumento2 pagineTAREA 5 BiocaRuth Milagros Ruiz RiosNessuna valutazione finora
- Práctica 8 Curvas de Crecimiento LogísticoDocumento3 paginePráctica 8 Curvas de Crecimiento LogísticoRuth Milagros Ruiz RiosNessuna valutazione finora
- Obj. 5 y 6Documento2 pagineObj. 5 y 6Ruth Milagros Ruiz RiosNessuna valutazione finora
- Preg.5 y 6Documento3 paginePreg.5 y 6Ruth Milagros Ruiz RiosNessuna valutazione finora
- INFORME 3 BiocaDocumento2 pagineINFORME 3 BiocaRuth Milagros Ruiz RiosNessuna valutazione finora
- Peces ExpoDocumento28 paginePeces ExpoRuth Milagros Ruiz RiosNessuna valutazione finora
- Cotos de CazaDocumento8 pagineCotos de CazaRuth Milagros Ruiz RiosNessuna valutazione finora
- Clase CHILOPODADocumento17 pagineClase CHILOPODARuth Milagros Ruiz Rios100% (1)
- Mapa Conceptual AdnDocumento1 paginaMapa Conceptual AdnDiego Iriarte50% (4)
- Cuadro Comparativo Linfocitos B y TDocumento6 pagineCuadro Comparativo Linfocitos B y TROGELIO LOPEZ LUEVANOSNessuna valutazione finora
- Introducción A La Endocrinología: Dr. Miguel Ángel Urquieta MaldonadoDocumento29 pagineIntroducción A La Endocrinología: Dr. Miguel Ángel Urquieta MaldonadoBarry AlenNessuna valutazione finora
- Serie MegacariociticaDocumento10 pagineSerie MegacariociticaAlexRendonNessuna valutazione finora
- Metabolismo de LipoproteínasDocumento1 paginaMetabolismo de LipoproteínasadeNessuna valutazione finora
- Bases Iónicas Del Aut. Cardiaco TMDocumento20 pagineBases Iónicas Del Aut. Cardiaco TMmatidialeNessuna valutazione finora
- Enzimas A BDocumento41 pagineEnzimas A BJlANessuna valutazione finora
- ?utf 8?b?r 3 Xdrwegzgugqw 5 HDG 9 TW 61 Hifbhdg 9 SW 7 Nnawnhlnbkzg ?Documento238 pagine?utf 8?b?r 3 Xdrwegzgugqw 5 HDG 9 TW 61 Hifbhdg 9 SW 7 Nnawnhlnbkzg ?Ale100% (1)
- Capítulos 1, 2, 3, 4, 5, 6, 7 y 8 de AnatomiaDocumento171 pagineCapítulos 1, 2, 3, 4, 5, 6, 7 y 8 de AnatomiaLeo C GNessuna valutazione finora
- CELULASDocumento4 pagineCELULASIMPRESIONESNessuna valutazione finora
- La Insulina y Sus Efectos MetabólicosDocumento7 pagineLa Insulina y Sus Efectos Metabólicoskatherin PCNessuna valutazione finora
- Final IiDocumento6 pagineFinal IiLetícia Raquel PintoNessuna valutazione finora
- 10egb Len Mat EESS CCNN F1 IntegradoDocumento3 pagine10egb Len Mat EESS CCNN F1 IntegradoKatherine Carranza BalderramoNessuna valutazione finora
- Serie MegacariociticaDocumento21 pagineSerie MegacariociticaDaliz WendyNessuna valutazione finora
- Fisión BinariaDocumento3 pagineFisión BinariakimNessuna valutazione finora
- Manual de SusceptibilidadDocumento248 pagineManual de Susceptibilidadhaeduga20004201Nessuna valutazione finora
- Remodelación ÓseaDocumento4 pagineRemodelación Óseadaniela restrepoNessuna valutazione finora
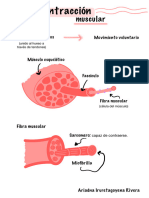
- Contracción MuscularDocumento3 pagineContracción MuscularAriadna IruretagoyenaNessuna valutazione finora
- Guia Semana 2-RESUELTODocumento7 pagineGuia Semana 2-RESUELTOClaudia CribilleroNessuna valutazione finora
- Publicacion PDFDocumento4 paginePublicacion PDFJimmy alexander gallego rodriguezNessuna valutazione finora
- Practica de Laboratorio La CelulaDocumento4 paginePractica de Laboratorio La CelulameidyreyezNessuna valutazione finora
- Taller de División CelularDocumento3 pagineTaller de División CelularSantiago GarciaNessuna valutazione finora
- 1 GTA CIENCIAS 8 Mario BarrantesDocumento19 pagine1 GTA CIENCIAS 8 Mario BarrantesMario BarrantesNessuna valutazione finora
- Regulación de La Síntesis de Galactosidasa En: Escherichia ColiDocumento53 pagineRegulación de La Síntesis de Galactosidasa En: Escherichia ColiZarate Arrieta Erika VianeyNessuna valutazione finora
- La Arginina Regula El Recambio de Proteínas en Las Células Epiteliales Mamarias Porcinas para Mejorar La Síntesis de Proteínas de La LecheDocumento8 pagineLa Arginina Regula El Recambio de Proteínas en Las Células Epiteliales Mamarias Porcinas para Mejorar La Síntesis de Proteínas de La LecheMARIAAAAAAAAAAANessuna valutazione finora
- 2do Parcial Histo 1 2020Documento36 pagine2do Parcial Histo 1 2020Ortellado Ramoa, Jorge RamónNessuna valutazione finora
- Haga Un Diagrama de Las Diferencias Entre El Extracelular y El IntracelularDocumento3 pagineHaga Un Diagrama de Las Diferencias Entre El Extracelular y El IntracelularYessenia BarrantesNessuna valutazione finora
- Exones y Intrones-Amino Acidos EsencialesDocumento2 pagineExones y Intrones-Amino Acidos EsencialesAnonymous 5yznLAxfrgNessuna valutazione finora
- Regulación AlostéricaDocumento5 pagineRegulación AlostéricaJosejuan DuranNessuna valutazione finora
- 120 PreguntasDocumento64 pagine120 PreguntasJudith CJNessuna valutazione finora