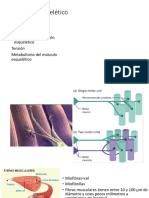
Potrebbero piacerti anche
- Apuntes Tejido MuscularDocumento18 pagineApuntes Tejido MuscularPaz GarciaNessuna valutazione finora
- Tejido MuscularDocumento7 pagineTejido MuscularShei SordianNessuna valutazione finora
- H06 Tejido Muscular Dra. Laura BarbozaDocumento8 pagineH06 Tejido Muscular Dra. Laura BarbozaPaolaNessuna valutazione finora
- Tema 4. - Tejido MuscularDocumento14 pagineTema 4. - Tejido MuscularFernanda Valeria Gutiérrez LeónNessuna valutazione finora
- Histologia de MusculosDocumento32 pagineHistologia de MusculosAndres CarrilloNessuna valutazione finora
- Ensayo de Fisiología DeportivaDocumento8 pagineEnsayo de Fisiología DeportivaDayana MorenoNessuna valutazione finora
- TEJIDO MUSCULAR - Ficha 02Documento4 pagineTEJIDO MUSCULAR - Ficha 02JOSUE LLASA PAUCCARANessuna valutazione finora
- Histología - MúsculosDocumento4 pagineHistología - MúsculosIgnacio N. AlmazánNessuna valutazione finora
- CLASE 7 FISIOLOGIA MUSCULARDocumento10 pagineCLASE 7 FISIOLOGIA MUSCULARKarlita Loncuante UrrutiaNessuna valutazione finora
- Tejido muscular esquelético: características y estructuraDocumento12 pagineTejido muscular esquelético: características y estructuraPierce PittyNessuna valutazione finora
- Estructra y Funciones Del Muscculo EsqueleticoDocumento13 pagineEstructra y Funciones Del Muscculo EsqueleticoMaricruz Cordova DuarteNessuna valutazione finora
- Histologia Tejido MuscularDocumento5 pagineHistologia Tejido MusculargomimdiegozamoragomimNessuna valutazione finora
- Tejido MuscularDocumento12 pagineTejido MuscularEduardo GálvezNessuna valutazione finora
- Seminario de Tejido MuscularDocumento19 pagineSeminario de Tejido Musculardanly castilloNessuna valutazione finora
- Tema 4-3Documento35 pagineTema 4-3MARIA DEL ROSARIO DUARTENessuna valutazione finora
- Tejido muscular: estructura y tiposDocumento16 pagineTejido muscular: estructura y tiposAbril CarinoNessuna valutazione finora
- Biomecanica FuerzaDocumento14 pagineBiomecanica FuerzaedielalondraNessuna valutazione finora
- Tejido MuscularDocumento4 pagineTejido MuscularJoyce ZambranoNessuna valutazione finora
- El Tejido MuscularDocumento9 pagineEl Tejido MuscularBeatriz GomezNessuna valutazione finora
- Músculos superficiales y medios: clasificación, estructura y funciónDocumento22 pagineMúsculos superficiales y medios: clasificación, estructura y funciónMelva Infanzon PomaNessuna valutazione finora
- Apuntes Tejido MuscularDocumento11 pagineApuntes Tejido Muscularedugalindo8Nessuna valutazione finora
- UNIDAD 02 (B) - Tejido MuscularDocumento7 pagineUNIDAD 02 (B) - Tejido MuscularJesúsNessuna valutazione finora
- Tejido MuscularDocumento37 pagineTejido MuscularKathya DiazNessuna valutazione finora
- Cap Tejido Muscular RossDocumento13 pagineCap Tejido Muscular RossnohemiNessuna valutazione finora
- Documento Completo - pdf-PDFA PDFDocumento18 pagineDocumento Completo - pdf-PDFA PDFoscarbio2009Nessuna valutazione finora
- Docsity Resumen de Histologia de Ross 1Documento27 pagineDocsity Resumen de Histologia de Ross 1Van NanaNessuna valutazione finora
- Tejido MuscularDocumento33 pagineTejido MuscularRocioNessuna valutazione finora
- Funcion de Los Musculos SuperficialesDocumento56 pagineFuncion de Los Musculos SuperficialesMaria Acosta100% (3)
- Fisiología de La MotricidadDocumento6 pagineFisiología de La MotricidadLuciana Napolitano100% (1)
- Sistema MuscularDocumento19 pagineSistema MuscularÁlex LomelíNessuna valutazione finora
- Sistema-Muscular Fisiologia FinalDocumento64 pagineSistema-Muscular Fisiologia Finaldanly castillo100% (2)
- Generalidades de Miología SekDocumento50 pagineGeneralidades de Miología SekArmandoRiosNessuna valutazione finora
- LocomacionDocumento7 pagineLocomacionCurso HD USM Sección ANessuna valutazione finora
- Contracción Muscular - GuytonDocumento9 pagineContracción Muscular - GuytonSanty LopezNessuna valutazione finora
- Manual Avanzado Capacitación en Elaboración de CárnicosDocumento149 pagineManual Avanzado Capacitación en Elaboración de CárnicosRolando GandolfoNessuna valutazione finora
- Tejido Muscular Material Complementario (Histología)Documento12 pagineTejido Muscular Material Complementario (Histología)miguel bravoNessuna valutazione finora
- RESUMEN FISIOLOGIA 1 ExamenDocumento101 pagineRESUMEN FISIOLOGIA 1 ExamenFavio Alexis DíazNessuna valutazione finora
- 2.3.1 Diferencias Histológicas Entre Fibras Musculares.Documento7 pagine2.3.1 Diferencias Histológicas Entre Fibras Musculares.joel garcia saldañaNessuna valutazione finora
- Tejido MuscularDocumento19 pagineTejido MuscularYohn LópezNessuna valutazione finora
- Generalidades Del Sistema MuscularDocumento6 pagineGeneralidades Del Sistema MuscularDaniela SantanaNessuna valutazione finora
- Sistema MuscularDocumento2 pagineSistema MuscularFiorella MoffaNessuna valutazione finora
- TEMA 2Documento11 pagineTEMA 2Ángela RodríguezNessuna valutazione finora
- Clases de MúsculosDocumento12 pagineClases de MúsculosMauroPogliacomiNessuna valutazione finora
- Tejido Muscular Estriado EsqueléticoDocumento20 pagineTejido Muscular Estriado EsqueléticoMatias HernandezNessuna valutazione finora
- Chopita - Tejido MuscularDocumento8 pagineChopita - Tejido MuscularIngrid CruzNessuna valutazione finora
- Tejido MuscularDocumento5 pagineTejido MuscularliloNessuna valutazione finora
- Tejido muscular: estructura, tipos y característicasDocumento4 pagineTejido muscular: estructura, tipos y característicasLorena MoreiraNessuna valutazione finora
- Sistema MuscularDocumento9 pagineSistema MuscularCarlos GaetanNessuna valutazione finora
- Biology Subject For High School Upper & Lower Arm Muscles by SlidesgoDocumento33 pagineBiology Subject For High School Upper & Lower Arm Muscles by Slidesgobykemy54Nessuna valutazione finora
- MiofibrillasDocumento4 pagineMiofibrillasItzel SolórzanoNessuna valutazione finora
- 5-Tejido MuscularDocumento7 pagine5-Tejido MuscularLuciana RodríguezNessuna valutazione finora
- Fisiología del músculo esquelético: funciones, propiedades y organizaciónDocumento11 pagineFisiología del músculo esquelético: funciones, propiedades y organizaciónInm GarciaNessuna valutazione finora
- Anatomia y Fisiologia Del Sistema MuscularDocumento22 pagineAnatomia y Fisiologia Del Sistema MuscularJ. Gabriel Torres67% (6)
- Tejido MuscularDocumento9 pagineTejido MuscularOscar PerezNessuna valutazione finora
- Sistema MuscularDocumento48 pagineSistema MuscularPaula GodinezNessuna valutazione finora
- Tejido muscular: clasificación y funcionesDocumento8 pagineTejido muscular: clasificación y funcionesDARWIN RUDY CAMPERO GUZMÁNNessuna valutazione finora
- Afi1 U2 Ea LaepDocumento7 pagineAfi1 U2 Ea Laeplaura EspinozaNessuna valutazione finora
- Tejido MuscularDocumento8 pagineTejido MuscularQuipyNessuna valutazione finora
- Histología y Embrología del Sistema NerviosoDa EverandHistología y Embrología del Sistema NerviosoValutazione: 3 su 5 stelle3/5 (1)
- Foro 14Documento3 pagineForo 14Christhopher TorresNessuna valutazione finora
- Aplicación de La Citometría A Flujo Al Diagnóstico de Las IDPDocumento2 pagineAplicación de La Citometría A Flujo Al Diagnóstico de Las IDPChristhopher TorresNessuna valutazione finora
- Inmunogenicidad de La Vacuna Contra HBV en NiñosDocumento3 pagineInmunogenicidad de La Vacuna Contra HBV en NiñosChristhopher TorresNessuna valutazione finora
- InmunoDocumento4 pagineInmunoChristhopher TorresNessuna valutazione finora
- Aplicación de La Citometría A Flujo Al Diagnóstico de Las IDPDocumento2 pagineAplicación de La Citometría A Flujo Al Diagnóstico de Las IDPChristhopher TorresNessuna valutazione finora
- Estrategias Sanitarias-1Documento12 pagineEstrategias Sanitarias-1Christhopher Torres0% (1)
- Medicina Tradicional-Informe2018Documento11 pagineMedicina Tradicional-Informe2018Christhopher TorresNessuna valutazione finora
- Estrategias Sanitarias II y IIIDocumento11 pagineEstrategias Sanitarias II y IIIChristhopher TorresNessuna valutazione finora
- Principios Básicos de ElectroterapiaDocumento44 paginePrincipios Básicos de ElectroterapiaAlejandro Sánchez PriegoNessuna valutazione finora
- Entregable 1Documento8 pagineEntregable 1LIZETH MARIA ORDOÑEZ RAMOSNessuna valutazione finora
- Test de IrwinDocumento75 pagineTest de IrwinYoselin Milagros Chambi Velasquez100% (2)
- Preguntes FISIODocumento8 paginePreguntes FISIOmargotNessuna valutazione finora
- Biomecánica Medica y BiodinámicaDocumento3 pagineBiomecánica Medica y BiodinámicaArianaNessuna valutazione finora
- Tarea de La NeuronaDocumento4 pagineTarea de La NeuronaJimmy Alexander Flores CanazasNessuna valutazione finora
- Guia 4 CompletaDocumento7 pagineGuia 4 CompletaDaniel Simón CruzNessuna valutazione finora
- Doctrina Naturipatica IiiDocumento26 pagineDoctrina Naturipatica Iiicar91% (11)
- Practica 1 Receptores SensorialesDocumento14 paginePractica 1 Receptores SensorialesEfrain CuellarNessuna valutazione finora
- Canales IonicosDocumento32 pagineCanales IonicosMario Leonardo Guevara EscalanteNessuna valutazione finora
- Planificación Clase A Clase 3º Medio BiologiaDocumento7 paginePlanificación Clase A Clase 3º Medio BiologiaJenny Castro Jara100% (1)
- Taller Evaluativo Anatomia y FisiologiaDocumento14 pagineTaller Evaluativo Anatomia y Fisiologiajimena cNessuna valutazione finora
- Practica HHDocumento10 paginePractica HHJaime MaldonadoNessuna valutazione finora
- Introduccion A La NeurofisiologiaDocumento15 pagineIntroduccion A La NeurofisiologiaFernanda CuadrosNessuna valutazione finora
- Potencial de AcciónDocumento16 paginePotencial de AcciónEsteban De AbreuNessuna valutazione finora
- Magazine 2018Documento100 pagineMagazine 2018some dude100% (2)
- Farmacología de Los Anestésicos LocalesDocumento2 pagineFarmacología de Los Anestésicos LocalesLuis Gustavo CárdenasNessuna valutazione finora
- Conduccion y Transmision Del Potencial de Accion Dra Oris L de Calvo MSCDocumento21 pagineConduccion y Transmision Del Potencial de Accion Dra Oris L de Calvo MSCMilagros RosasNessuna valutazione finora
- Atencion Al Paciente PolitraumatizadoDocumento15 pagineAtencion Al Paciente PolitraumatizadoJuan Carlos Pérez MartínezNessuna valutazione finora
- Resumen Semestral de BiologiaDocumento52 pagineResumen Semestral de BiologiaMario HerreraNessuna valutazione finora
- Potencial de Reposo de Membrana y Potencial de Acción PDFDocumento12 paginePotencial de Reposo de Membrana y Potencial de Acción PDFJuan Jose CalvacheNessuna valutazione finora
- Solemne 1 FisiologíaDocumento18 pagineSolemne 1 FisiologíaMatias AstudilloNessuna valutazione finora
- Tejido NerviosoDocumento7 pagineTejido NerviosoLeandro Sebastián Guzmán BuchelliNessuna valutazione finora
- SinapsisDocumento4 pagineSinapsisCesar Duque50% (2)
- Types of Waves Science Presentation in Light Blue White Lined StyleDocumento13 pagineTypes of Waves Science Presentation in Light Blue White Lined StyleCatalina CelisNessuna valutazione finora
- Fisiología General de Los ReceptoresDocumento6 pagineFisiología General de Los ReceptoresAngel Luis Alvarez GonzálezNessuna valutazione finora
- Seminario 9 - Práctica 9 - Musculo CardiacoDocumento14 pagineSeminario 9 - Práctica 9 - Musculo CardiacoMarcos MoralesNessuna valutazione finora
- Informe de Fisiologia 2Documento6 pagineInforme de Fisiologia 2Marcos MontalvanNessuna valutazione finora
- MúscularDocumento45 pagineMúscularAxel EstradaNessuna valutazione finora
- Neuro Modulo KenedyDocumento35 pagineNeuro Modulo Kenedymaria stafNessuna valutazione finora