Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
CAPÍTULO - 13-Tejido Muscular PDF
Caricato da
LauraSeoaneIvanacTitolo originale
Copyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
Formati disponibili
CAPÍTULO - 13-Tejido Muscular PDF
Caricato da
LauraSeoaneIvanacCopyright:
Formati disponibili
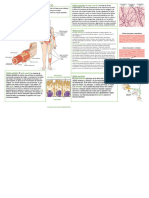
Tejido muscular
"En ocasiones las lÍneas de un diseño coIIen en ditección opuesta a lo esperado.
Sin embargo, sigue siendo ttn diseño."
Karen Blixen
El movimienfo activo orientado es ca- vísceras flat. viscus, pl. viscera) y es iner-
r a c t e r í s t i cdoe l a s f o r m a sv i v a ss u p e r i o r e s vado por el sistema nervioso autónomo
y los organismosmulticelulares han desa- (involuntario). Por Io tanto, a menudo la
rrollado células muy especializadas, las musculatura lisa se denomina musculatu'
células musculares,para salvaguardarIas ra visceral o involunLorio.
necesidades de movilidad internay exter- El músculo esquelético está compuesto
na que tiene ei organismo. por células muy largas, cada una de las
Las células musculares son alargadas, cuaies contiene gran cantidad de núcleos
con el eje longitr.rdinal orientadoen la di- ubicados en la periferia (fig. 13-1). Todos
iección del movimiento,a menudo en for- los músculos del movimiento están for-
ma tan expresaque antesse las denomina- mados por mÍrsculo esquelético. Las célu-
ba fibras. EI término fibra muscular aún las presentan un estriado característico,
se emplea,si bien se trata de células,a di- por'lo qrie la musculatura esquelética
ferencia de las fibras extracelularesde te- también se denomina musculatura estria-
'iido conectivo. do. Otra denominación es musculaturo
En el organismo de los vertebrados voluntaria, como consecuencia de la iner-
e¡isten tres tipos de musculaturabien di- vación oor el sistema nervioso somático
ferenciadas por estructura y función: (voluntário).
músculoliso, múscttloesqueléticoy mús- El músculo cardíaco está compuesto
culo cardíaco,con propiedadesgenerales por célnlas con núcleo central, como el
- se detallana continuación.
oue múrsculo liso, pero con estriado transver-
El músculo liso estácompttestopor cé- sal similar al de Ia musculatura esqueléti-
lulas ahusadas.cada Llna con su nÍtcleo ca (fig. 13-1). La musculatura cardíaca só-
central (fig. 13-1).El músculo liso se en- lo se encuentra en el corazón y es inerva-
cuentra,por ejemplo.en las paredesde las da por el sistema nervioso alttónomo.
ri
Fig. 13-1. Dibujosesque- Músculoliso Músculoesquelético Músculocardíaco
máticosque muestranlas
características histoló-
gicas básicas de los
tres tipos de músculo,
como se ven en cortes
longitudinales (parte su-
perior) y transversales
(parte inferior), respecti-
vamente,con el mlcrosco-
pio óptico.(SegúnLeon-
hardt.)
CAPITULO tó TEJIDO MUSCULAR 299
Fig. 13-2. Dib
Músculo liso máticode fibr¿
lares lisas ais
La musculatura iisa se encuentra en ca-
de maceració
si todos Ios órganos y vasos y desempeña nítrico"
un papel importante en la función de la
mayoría de los órganos y de los sistemas
de órganos.
Características del músculo liso
con el microscopio óptico
Las fibras musculares lisas son células
largas ahusadas, es decir con extremos afi- mita con los extremos afinados de las cé-
lados (fig. 13-2). Pueden aparecer aisladas, lulas adyacentes. En consecuencia, en un
pero se suelen agrupar en capas donde es corte transversal hay notable variación
difícil determinar los límites de las células del diámetro del contorno de las fibras
individuales con el microscopio óptico. El seccionadas (fig. 13- ) y sólo se observan
tamaño de las fibras es muy variabie. Las cortes transversalesdel nricleo en los con-
fibras más grandes se encuentran en el úte- tornos más grandes.
ro grávido-(hasta 10 x 500 pm), mientras El citoplasma de las fib¡as musculares
que las más pequeñas se encuentran en las lisas y estiiadas se denomina sarcoplasma
arteriolas (Z x rs ¡rm). (gr. sorx, carne). En los cortes coloreados
Cada fibra muscular posee un úrnico con hematoxilina-eosina el citoolasma se
núcleo localizado en la oorción media l i ñ e d e r o j o p o r l a e o s i n a .E l c o i o r e s r o j o
m á s a n c h a ( t i g s .1 3 - 2 y l 3 - 3 ) y e n l o s c o r - más intenso y más violeta que en la colo-
tes transversaies aparece en e1 centro (fig. ración de las fibras de coláeeno. También
1 3 - 4 ) .E l n ú c l e o e s a l a r g a d oe n e l s e n t i d o c o n t r i b u v e a l a d i [ e r e n c i a c i é ne n t r e l a s c é -
longitudinal de Ia fibra y posee extremos lulas musculares y las fibras de colágeno
afinados o redondeados. A menudo en la tinción mediante métodos más selecti-
los cortes histológicos se observan incur- vos, como por ejemplo el de van Gieson,
vaciones o arrollamientos del núcleo. Se que colorea de rojo las fibras de colágeno y
piensa que son artificios. La cromatina de amarillo el sarcoplasma (fig. 13-5).
suele ser periférica y se distinguen varios Mediante tinciones especialesse puede
nucléoios. demostrar Ia presencia de las organelas
Las células de los haces o caoas están habituales del citoplasma y, en algunos
d e n s a m e n l ee m p a q r r e t a d a sp.o r l o q u e l a casos, de glucógeno. Las organelas se r.rbi-
porción media más ancha de una célula li- can, en su mayor parte, cerca de los poios
nucleares.
Las capas o haces de fibras musculares
Fig. 13-3. FotomicrografÍa de un corte longitu- lisas se mantienen unidas oor medio de
dinal a través de células musculareslisas de l e j i d o c o n e c t i v o .d e s d ed o n d e p e n e t r a nf i -
la pareddel intest¡nogrueso Cortecoloreado nas fibras de tejido conectivo entre las fi-
con hematoxilina-eos¡na. x440 bras musculares y allí, por último, una
Células
muscularesilsas
Fig. 13-4. Foto
fía de un corte
sal a través de
musculareslis
pareddel intes
;:,. so. Corte color
* s,' hematox¡l¡na
:$ x 1. 1 0 0 .
3OO TEJIDO MUSCULAR CAPITU,
Fig. 13-5. Fotomicrogra- Células musculares lisas Núcleos
fía de un corte transver-
sal a través de células
musculares lisas de la
pared del tubo digestivo.
Corte coloreadopor el
métodode van Gieson.
x540.
delgadared de fibras reticularesrodea ca-
da célula muscular. Estas se impregnan
con plata y también con Ia tinción de PAS
se demuestraIa presenciade una capa de
glucosaminoglucanossobre Ia superficie
de cada fibra muscular. Esta capa es una
lámina externa y la red de fibras reticula-
res corresDondea la lámina reticular de Ia
membraná basal (véase membrana basal
en el cap. 6, pág.169).Durantela contrac-
ción se transmite Ia contracción de las fi-
bras muscularesindividualesa la capade GlucógenoCondensación
teiido conectivo circundante a travésde Ia
vaina de fibras reticulares. Fig.13-6.lmagende un cortelongitudinal a
travésde la partecentralde una célulamus-
Ultraestructura de la musculatura lisa cular lisa,captadacon microscopio electrónico.
Nóteseel sarcoolasma librede filamentosco-
Con el microscopioelectrónicose obser- rrespondiente al polonuclear,donde,entre
se distinguenmitocondriasy
va una zona de sarcoplasmalibre de fila- otrasorganelas,
variosgránulos de glucógeno. En el sarcoplas-
mentosen los polos nuclea¡esque contiene ma periférico se observan miofilamentosagru-
mitocondriasaisladas,un pequeñocomple- padosquetranscurren en direcciónlongitudinal,
jo de Golgi y escasosperfilesde retÍculoen- ademásde condensaciones citoplasmáticas
doplasmáticorugoso(fig. 13-6).Hay retícu- dispersas. En el espaciointercelularse obser-
lo endoplasmáticoliso bajo la forma de ele- vanfinasmicrofibrillas de colágeno. x22.0O0.
mentos tubulares de transcurso longitudi- (Cedida porJ.V Small.)
nal ubicadosentrelos filamentosy se deno-
minq retíeqlg qqlaoplqslllático.
Cerca de la superficie citoplasmática ma. oue se ha demostrado contienen Ia
del plasmalemase distinguennumerosas proteina fijadora de actina alfaactinina.
cavéolas similares a vesículas de pinoci- Además,a menudo el plasmalemaestáen
tosis en proceso de liberación que no se contacto con las células muscularesveci-
concreta. Sobre Ia superficie interna del nas mediante nexos típicos. En los sitios
plasmalema, entre las cavéolas,también en las que las células no estánunidas por
se observan?onas electrondensasdisper- nexos hay un espaciode 50-80 nm de es-
sas o placas de inserción.(fig.13-7),con pesorocupadopor los componentesde la
característicasde adhesionesfocales, da- lámina extracelular.También se observan
do oue se ha demostradomediante méto- fibras elásticas aisladas producidas, al
doslnmunohistoquímicos la presenciade igual que los demás componentes de la
tallina y vinculina, que contribuyen a fijar matriz, por las células musculareslisas-.
los filamentos de actina a la placa (véase parte {el sqrcoplasmaes ocu-
.!a ¡¡r.q)¿or
adhesionesfocalesen el cap. 6, pág.1,64, pado por fi_la¡nenlqs,en p-ar19delgados fi-
y la fig. 6-13).Del mismo modo se encuen- lamentos de actina y gruesos filament-o-s-,..
tran condensacionescitoplasmáticasa in- de miosina y en parte filamentos interme-
tervalos regulares por todo el sarcoplas- üos (fig. r3-7).
CAPITULO 13 TEJIDO MUSCULAR 301
Los delgados ül4mentos de acJina_tie- Filamentos (en la condensación)
intermedios
nen un diámetro de unos 7 nm. Los fila-
mentos de actina de las células muscula-
res, incluso las células musculares lisas,
son del tipo estable, que no se confunde
con los monómeros de actina G en un
pool soluble (véase cap. 3, pág. 91).
Los gruesos filamentos de miosina tie-
nen, en promedio, unos 15 nm de diámetro.
En las imágenes captadas con micros-
copio electrónico, cada filamento de mio-
sina se caracteriza por estar rodeado por
un anillo de delgados filamentos de actina
(fig. 13-7), de modo que Ia relación total
entre filamentos de miosina y filamentos
de actina es de 1:15.
Los filamentos de actina y de miosina
se reúnen en Brupos o unidades. Estas
unidades o haces se extienden desde una
condensación citoolasmática hasta la si-
guiente, dado que él haz se fija a alfaacti-
nina en la condensación (fig. 13-B). Los fi-
Iamentos de actina se unen con sus exrre-
mos positivos a la condensación, del mis-
mo modo como ocurre en la musculatura
estriada, por lo que se puede considerar a
la condensación como la contrapartida
funcional de Ias líneas Z y, al igual que és- Filamentos
de miosina Filamentos
de actina
tas, también contienen alfaactinina (véase
más adelanteJ. Así, los haces de filamen-
tos se pueden considerar unidades con- Fig. 13-7. Imagende parte de un corte trans-
tráctilei que, como una especie de sarcó- versal de una célula muscular lisa, captada
mero de oreanización laxa se extiende en- con microscopioelectrónico.Nóteseque los
gruesosfilamentosde miosinaestán rodeados
tre dos zoñas condensadas. Algunos de
por un anillode finosfilamentosde actinay que
los haces de filamentos también se unen a
los filamentosintermedios sólo se encuentran
las placas de inserción sobre Ia superficie en las condensaciones del citoplasmay en las
interna del plasmalema. placasde inserciónsubsarcolémicas. x100 000.
Los filamentos intermedios tienen un ( S e g ú nS m a l l . )
diámetro de unos 10 nm v están comDues-
tos por desmina (en las télulas muscula-
res lisas de las paredes vasculares están
formados pot vimentina). Los haces de fi- tienen distinta polaridarl, es decir, se
lamentos intermedios se extienden entre orientan en direcciones opuestas (véase
las condensaciones citoolasmáticas v en- con más detalle cabezasde miosina y des-
t r e é s t a sy l a s p l a c a s d e i n s e r c i ó n , y c o n - lizamiento de filamentos bajo muscr.rlatu-
forman un fuerte citoesqueleto en ei cuai ra esquelética). En consecuencia, durante
también intervienen filamentos de actina el deslizamiento. uno de ios filamentos de
distintos del tioo muscular. miosina puede "traccionar" de los fila-
Mecanismo de contracción en las célu- mentos de actina, filados cada uno a su
las musculares lisas. No está tan bien de- condensación citoplasmática, por lo que
finido como el de Ia musculatura estriada, éstas se acercan (fig. 13-B). AsÍ, el efecto
pero parece corresponder a éste en lo conjunto sobre toda la célula es el acorta-
principal, es decir, es un meconismo de miento, dado que la tracción también es
deslizamiento en el que los filamentos ri- transferida a las placas de inserción por
cos en actina y en miosina representan la intermedio del citoesoueleto. La muscu-
base estructurol (fig. 13-B). _A diferencia latura lisa se puede acortar en más del
de ia musculatura estriada, los filamentos B0%, es decir, mucho más que la muscu-
de miosina de las células musculares lisas latura estriada (unos 4 ¡rm contra alrede-
no poseen una zona media sin presencia dor de 1 pm), lo cual permite un mayor es-
de puentes transversales, es decir, sin ca- tiramiento para el deslizamiento de los fi-
bezas de miosina; éstas se encuentran en lamentos y una migración consecuente
toda la longitud del filamento de miosina. más prolongada de los filamentos de acti-
Por otra parte. Ios filamentos de miosina na opuestos. Además, los filamentos de
son más áplanados y se cree que las cabe- actina tienen la posibilidad de desplazar-
zas de miosina de las dos "superficies" se a lo larso de toda la extensión del fila-
302 TEJIDO MUSCULAB CAPiTU
mento de miosina, debido a la polariza- iones calcio. Después de la separación de
ción diferente, sin una zona central sin ca- los grupos fosfato la molécula de miosina
bezas de miosina. ya no posee la capacidad para fijarse a Ia
L.,3,pontracción muscular se inicia cqa¡; actina, se interrumpe la contracción y se
do la concentración de iones calcio att- relaja Ia célula muscular. Esta fg4q!"lt
menta en el citosol por difusión de iones -gulqci{n de la cqntlqce i-ónmuscular se de-
calcio hacia el interior de la célula desde nomina regulación ligada a miosina.
el espacio extracelular o salida del retícu- Algunos tipos de células musculares li-
lo sarcoplasmático, que conti-ene un depG sas también tienen un mecanismo de re-
qi!-o _de_ee]_ciO... L a .¡nay or c o nc entrac i ó n d e gulación supletorio, denominado regula-
iones calcio implica su unión con la ca-1- ción ligada a actina. Aquí el aumento de
modulina, y el complejo Ca**-calmodulina la concentración de iones calcio, con for-
activa entonces la enzima quinasa de ca- mación del complejo Ca**-calmodulina
denas livianas de miosina (MClquinasa) produce la activación de la proteína cal-
(ing. myosin light chain quinase). La MCL- desmona, una proteína reguladora unida
ouinasa cataliza la fosforilación de la cade- a actina, que impide la unión con miosi-
na liviana reguladora de la cabeza de mio- na. No obstante, la activación de la caldes-
sina, Io cual desencadena un cambio de mona causa un cambio en su conforma-
conformación de la molécula de miosina, ción, por lo que se separa de la actina y
que entonces es capaz de fijarse a la acti- oermite el acceso a Ia miosina. De este
na, por lo que comienza la contracción modo se realiza la unión cruzada entre ac-
(véase más adelante bajo mecanismo de tina y miosina y se inicia Ia contracción.
contracción de la musculatura esqueléti- Algunas células musculares lisas poseen
c a ) . C u a n d o l a c o n c e n t ¡ a c i ó nd e i o n e s c a l - otro mecanismo equivalente, pero con Ia
cio nuevamente desciende en el citosol se proteína calponina como reguladora en
inactiva la MClquinasa y al mismo tiempo Iugar de la caldesmona.
se escinde fosfato de Ia cadena liviana de La regulación ligada a miosina de la
miosina reguladora, proceso catalizado contracción, a veces complementada por
oor la enzima fosfatasa de cadenas livia- la regulación ligada a actina está amplia-
nas de miosina que está siempre activa mente difundida en céluias no muscula-
con independencia de la concentración de res en las que tiene lugar la contracción
dependiente de actina o miosina, por
ejemplo en la formación de la hendidura
de escisión relacionada con la división
Fig. 13-8. Dibujoesquemático de la posible del citoolasma durante la división celular.
organización del aparato contráctil y del ci- El ingreso de iones calcio al citosol
toesqueletoen las células musculareslisas (desde el espacio extracelular y/o los de-
(véaseel texto para los detalles)
pósitos del retículo sarcoplasmático) que
desencadena la contracción se puede ge-
nerar debido a varias acciones diferentes
Placade inserción sobre la célula muscular lisa. En este sen-
Filamento
de actina tido, esta céluia demuestra mayor varia-
ción que la célula muscular estriada, cuya
principal fuente de excitación son los im-
oulsos nerviosos. Esto se debe considerar
én relación con las numerosas funciones
Filamento importantes diferentes para Ia regulación
de miosina del medio interno del organismo. Además
de la estimulación nerviosa (véasemás so-
bre inervación de la musculatura lisa bajo
Unidadde
filamentos histofisiología), es decir la acción de los
neurotransmisores, las células musculares
f.isaspueden ser estimuladas para realizar
Corte longitudinalde Ia contracción oor hormonas circulantes o
Condensación una unidad de filamentos moléculas señál locales (paracrinas), por
citoplasmática estiramiento. oor modificaciones del con-
tenido de metabolitos del líquido extrace-
lular circundante y, por último por activi-
dad eléctrica esoontánea del sarcolema.
La contracción finaliza como conse-
cuencia de 1a disminución de la concen-
tración intracelular de iones calcio y se
debe a Ia eliminación de éstos por recap-
Corte transversal de una tación al retículo sarcoplasmático, media-
unidad de filamentos
do por una ATPasa activada por calcio,
CAPíTULO Ió TEJIDO MUSCULAB 303
Acoplamiento excitación-contracciónelectromecánicoy farmacomecánico
en las células musculares lisas.
En general, los mecanismos que aco- so de iones calcio al citosol desde el es-
olan la excitación de la célula muscular pacio
- extracelular.
lisa con la contracción se clasifican en El acoplamiento farmacomecánico no
acoplamientos electromecónicos y far- depende de la despolarizacióndel plas-
macomecónicos. malema, dado que implica la activación
El acoplamiento electromecánicoim- por unión de ligandos específicosa los
plica variacionesdel potencial de mem- canales de calcio activados por rccepto-
brana con despolarizacióny creación de ¡es, que se abren, por ejemplo después
un potencial de acción.Durantela despo- de la unión de un neurotransmisor(ca-
Iarizacióntiene lugar un ingresode iones nales de calcio dirigidos por transmiso-
calcio a Ia célula a través de canalesde res) o por una hormona.Además de estos
calcio dirigidos por potenciales (véase receptoresacopladosa los canalesióni-
membrana celular en el cap. 3, pág. 52). cos (véasecap.7, pág. 1,92)el transmisor
La despolarización puede ser causada o la hormona se puede unir a los rccep-
por varias de las acciones descritas,in- tores acoplados a Ia proteína G con li-
iluso la unión de un neurotransmisora beración de IP" (trifosfato de inositol),
un canal de calcio dirigido por potencia- que despuésde su formación en el cito-
les. Los canalesde sodio tienen menor plasma difunde al retículo sarcoplasmá-
importancia en Ia despolarizaciónde las tico, donde facilita Ia Iiberación de io-
células musculareslisas, dado que el po- nes calcio (véasecap. 7, pá9.192).La li-
tencial de membrana de -50 milivolt beraciónde iones calcio desdeel retícu-
(comparado con los -70 milivolt de la Io sarcoplasmático también se puede
musculatura estriaday los nervios) inac- producir por despolarizacióndel plas-
tiva los canales de sodio dirigidos por malema o por Ia denominada Liberución
ootenciales. También el estiramiento de iones calcio inducida por calcio, de-
puede causar la despolarización. Otra bida a que los iones calcio que ingresa-
causaes la transmisión desdeuna célula ron a la célula estimulan la ulterior li-
vecina a travésde nexos.En todos los ca- beración de iones calcio por el retículo
sos el resultado es el aumento del insre- sarcoplasmático.
además de Ia extracción desde la célula ción es rápida y es seguida por relajación
por bombeo a través de Ia correspondien- comoleta. se denomina contracción fási-
te bomba en el olasmalema. Una cantidad ca, Las fibras nunca muestran contraccio-
menor de iones-calcio es extraída de la cé- nes espontáneas. Este tipo se encuentra,
Iula a través de un mecanismo de inter- por ejemplo en el iris del ojo (véase con
cambio de sodio-potasio, por el cual tres más detaile en el cap. 24), el conducto de-
iones de sodio del esoacio extracelular ferente y los vasos de mayor calibre.
son intercambiados poi un ion de calcio El tipo unitario o visceral está com-
del citosol (la energía es provista por el puesto por densos haces o capas de célu-
gradiente de sodio en el plasmaiema, por las musculares unidas por nexos. Este ti-
lo que depende de la bomba de Na*-K* del po se caracteriza por la capacidad de con-
plasmalema). Este meanismo tiene impor- traerse espontóneamenfe, por ejemplo,
tancia mucho mayor en la musculatura oor la extensión más allá de cierto límite.
cardíaca (véase más adelante). Esta reacción ante el estiramiento tiene
gran importancia, porque causa el vacia-
'K Inervación de la musculatura lisa miento de un órgano hueco cuando la luz
se exoande hasta alcanzar un valor um-
Las propiedades de la musculatura lisa bral. Las contracciones espontáneas se di-
varían mucho de un órgano a otro, pero en funden o trovés de ]os nexos a las fibras
general se habla de dos tipos principales: vecinas, por lo que se genera una activi-
eI ttpp multiunitario y el tipo visceral o dad progresiva. La inervación es abun-
unitario. dante, pero el efecto de Ia acción nerviosa
El tipo multiunitario se compone de fi- está dirigido sobre todo a modificar la ac-
bras unitarias oue funcionan con inde- tividad espontánea. La velocidad de la
pendencia entre sí y que a menudo son contracción es baja y se mantiene una
inervadas por una única terminación ner- contracción constante prolongada, deno-
viosa. La activación tiene lugar por difu- minada c-ontracción tónica o tono. El tipo
siOn de un potencial de acción, Iácontrac- unitario se encuentra en las paredes de la
304 TEJIDO MUSCULAB CAPITU
mayor parte de los órganos, por ejemplo, musculareslisas la contracción es estimu-
el tubo digestivo, las vías biliares, las vías lada por la inervación adrenérgicae inhi-
urinarias y el útero, de allí Ia denomina- bida por Ia colinérgica,mientras en otras
ción tipo visceral. Las contracciones rít- ocurre lo contrario.Esto se debea diferen-
micas pueden ser cubiertas por un estado tes subtipos de receptorespara la misma
de contracción tónica. La musculatura li- sustanciatransmisora(véaseel cuadro so-
sa de las oaredes arteriolares está contraí- bre mecanismode acción de las proteínas
da en forma tónica con cierto grado de va- G , e n e l c a p . 7 ,p á g .1 9 0 ) .
soconstricción, por lo que se mantiene la
oresión arterial en determinado nivel. Histogénesis de la musculatura lisa
Áquí, las células musculares lisas poseen
receptores para hormonas circulantes que Las células musculareslisas se desarro-
actúan sobre el grado de contracción y, en llan a partir de células mesenquimáticas
consecuencia sobre el flujo sanguíneo y la embrionarias(sonuna excepciónlos mús-
oresión arterial. entre ellas Ia noradrenali- culos del iris, de origen neuroectodérmi-_
ña y Ia angiotensina, que causal vaso-, co). Durante la diferenciaciónse prolon-
constricción, y la bradiquinina y las pros- gan las células y surgenlos miofilamentos
taglandinas, que causan vasodilatación. El en el citopiasma,es decir, aparecenlos
tipo unitario se caracteriza por su elevado mioblastos,que sufren mitosis y se obser-
grado de regulación por acción hormonal, va una diferencición gradual a células
en contraDosición con la inervación. La musculareslisas. Se ha demostradoque
a c c i ó n d e l o s e s t r ó g e n o sc a u s a c r e c i m i e n - incluso célulasmusculareslisastotalmén-
to en tamaño de las fibras musculares del te diferenciadas poseen Ia capacidad de
útero durante el embarazo y la musculatu- sufrir mitosis durante toda Ia vida. pero es
ra uterina también es sensible a la oxitoci- raro observaruna mitosis en un ádrlto.
na, en especial en el periodo perinatal Sin embargo,hay aumento en tamañoy en
lvéanse con mavor detalle las hormonas cantidad de células musculareslisas, por
en los caps. 21 y 22). ejemplo,_eng! g!q1o durante el embarazo
La inervación de la musculatura lisa y ademásson comuneslas mitosis en las
tiene Iugar a través de las vías simpáticas qélutasmusculareslisas de las paredesde
y parasimpáticas del sistema nervioso los vasos.posiblementecomo un eslabón
autónomo (véase cap. 14). Por inervación en la renovaciónconstantede Ias células
de las células musculares lisas del fipo lesionadaso enveiecidas.
mulliunitario cada axón oosee varias ia-
mificaciones que se extiénden entre las
fibras de músculo liso. En los sitios de Músculo esquelético-t
contacto entre las ramificaciones y las fi-
bras musculares se interrumoe la vaina La fibra es la mínima unidad estructu-
de Schwann y el axón desnu-do crea ex- ral y funcional del músculo esquelético.
pansiones en los puntos de contacto, las Es una célula largar4ultinucleadaque se
varicosidades, que contienen la sustancia puede observarcon el microscopio ópti-
transmisora (acetilcolina o noradrenali- co y, en algunos casos,incluso a simple
na). En la inervación de las células mus- vista. Las fibras muscularesse reúnen en
culares lisas del tipo unitario las ramifica- haceso fascículos (Iai. fascis, haz), que a
ciones del axón nunca entran en contacto su vez forman los distintos-tiposmuscu-
directo con las fibras musculares lisas. lares (fig. 13-9ay 13-9b).Un músculo es-
Las ramificaciones se enroscan entre las tá rodeadopor una capa de tejido conec-
fibras o llegan hasta el tejido conectivo t i v o , e l e p i m i s i o ,q u e p o r s u p a r t e e s t á
que rodea los haces o las capas de fibras. más o mños entr;tejido con-la loscio
Las ramificaciones del axón también aouí muscular circundante fvéasecap. B, pag.
forman varicosidades y la sustancia trañs- 223). El epimisio se extiende hacia el in-
misora liberada debe difundir a través del terior del músculo y rodea todos Ios fas-
espacio extracelular, a menudo por dis- cículos como perimisio (fig. 13-10),que
tancias considerables, para llegar a las fi- por último forma una delgada vaina de
bras musculares inervadas. Las células fibras reticulares, el endomisio, alrede-
musculares lisas poseen receptores coli- dor de cada fibra muscular. funto con los
nérgicos del tipo musca¡ínico (en aparien- glucosaminoglucanos,las fibras reticula-
cia éstas están localizadas en las células res contribuyen a la formación de una Jd-
endoteliales, que afectan indirectamente mina externo alrededor de cada fibra
las células musculares lisas, véase cap. muscular, del mismo modo que alrede-
1,4,pág.341), sensibles a acetilcolina, ade- dor de las células musculareslisas. Los
más de receptores alfaadrenérgicos y be- músculos están fijados al esqueleto me-
taadrenérgicos para noradrenalina. Estos diante tendonesunidos al epimisio.Ade-
últimos también son activados por la más de unir las fibras y los fascículos
adrenalina circulante. En algunascélulas musculares,Ias vainas de teiido conecti-
CAPíTULO 13 TEJIDO MUSCULAR 305
Fig. 13-9. Dib
máticode la c
ción del mús
quelético des
macroscópic
cular. (Según
Fawcett.)
Fascículo muscular
Fibra muscular
Banda Línea Banda Banda-
H z A
E
Miofibrilla Sarcómero
Moléculasde actinaG
H t ttt!
e
[--
I ---
Miofilamentos
|
I ---
|
Filamentode actinaF
Filamentode miosina
Moléculade miosina
i "..
oao
aoaa n
aoaaa
ooaa o.'oio!o Meromiosina Meromiosina
aaa ¡'r"o liviana pesada
vo permiten a cada fibra y fascículo el humano, la longitud promedio de la fibra
movimiento independiente. varía desdemenos de 1 mm (músculo del
Los.vasossanguíneosy los nervios atra- estribo)hastamás de 30 cm (músculo sar-
viesan las vainas de tejido conectivoy pe- torio). En la mayoría de los músculos las
netran en el interior del músculo, y cada fibras son más cortasque toda la longitud
fibra muscular está rodeadaoor uña rica del músculo y se fijan a tabiquesde tejido
¡qd qapilqr. conectivo musculares por el extremo no
f i i a d oa t e n d ó n .
Característicasdel músculo estriado El diómetro de Ia fibra varÍa entre 10 y
con el microscopio óptico 100 pm. En un músculo determinados-e
encuentrauna mezcla de fibras de distinto
Las fibras del músculo esoueléticoson espesor,pero de todos modos hay un espe-
u n a n o t a b l ev a r i a - sor oromedio característicode las fibras.
c i l í n d r i c a sv o r e s e n t a n
ción de longiiud y de diámetro. En el ser El aumentodel tamaño de un músculo du-
306 TEJIDOMUSCULAR CAPíTU
vantador de pesas). El aumento de espqso:
Epimisio por entrenaÁiento se denomina hipérto-
fiq (gr. fro/e. nutrición. crecimienl.o),
mientras que la disminución del espesor
de las fibras y la consiguiente reducción
dé la masa muscular por falta de uso, por
ejemplo, en caso de fractura, se denomina
atrofia.
En preparados macerados y extraídos
por punción y analizados con microsco-
pio óptico se observa que cada fibra está
rodeada por una delgada membrana trans-
parente, de alrededor de 1 pm de espesor,
formada por la unión del plasmalema y la
lámina externa (fig. 13-11). -No es común
observarla en los cortes histológicos habi-
Fibramuscular
tuales. En las células musculares el plas-
malema se denomina sarcolema.
Gada fibra muscular contiene finas fi-
Fig. 13-10. Dibujoesque- rante el desarrollodel individuo o por en- brillas estriadas paralelas, denominadas
málicode las distintas trenamientose debe a incrementodel es- miofibrillas, por lo general de un espesor
partes de tejido conecti- pesor de cada fibra (cf. las imágenesde la d e 1 - 2 u m f f i s . 1 3 - 1 2 ] .E n l o s c o r t e s t r a n s -
vo que intervienen en la izquierda y de la derechade la fig. 13-29, versales a menudo se observa que están
conformación de un que muestranfibras de músculo esqueléti- agrupadas en los campos de Cohnheim
músculo esquelético.
(SegúnGrollman.) co de una personano entrenaday de un Ie- (fig. 13-11), un artificio. El est-riadq trans-
velsal se debe a que cada miofibrila está
formada por segmentos alternados con re-.
gularidad, de diferente índice de refrac-
-c¡{¡r, Toda Ia fibra aparece estriada, por-
que en las miofibrillas paralelas la misma
Miofibrillas(1-2pm) clase de segmentos se ordena a la par (figs.
13-1.2y 13-14).
ta fibra de músculo esquelético o me-
Núcleosde la
fibramuscular nudo contiene varios cientos de núcleos,
localizados justo por debaio del sarcolema
(fig. 13-1). La ubicación periférica de los
núcleos se distingue mejor en los cortes
Fibra muscularrota
Sarcolema
Fig. 13-12. FotomicrografÍa del músculo es-
Célulasde tejidoconectivo quelético. Se observauna parte de dos fibras
musculares,separadaspor un angostoendomi-
sio (en el que se visualizaun capilar).Con clari-
Extremosromosde la fibra dad se distingueque las fibras musculareses-
tán compuestaspor miofibrillascon estrías
transversales. Corte de una muestraincluidaen
plásticoEpon coloreadacon azul de metileno.
x2.2O0.
Miofibrillas
Camposde
Cohnheim
Sarcolema
Fig. 13-11. Dibujoesque-
máticode la conforma-
ción detallada al micros-
copio óptico de las fi-
bras del músculo esque-
lético. (SegúnBródel.) BandaA Banda I LíneaZ Banda H
CAPITULO 13 TEJIDO MUSCULAR 307
(fig. 13-13). En los cortes Núcleos Fig. 13-13. F
!¡g¡rqyerqqle-s fía de un corl
longitudinales algunos núcleos p¿rrecen
sal a través r
estar ubicados en la parte media de la fi- musculares r
bra, pero en realidad estánen la superficie Corte colorea
o en la profundidad, de acuerdocon el es- matoxilina-e
pesor de la muestra.Los _núcleosson apla-
iados y ovales en el seiti-ilóTongtnü-aaf
de la fibra y están dispersosa lo largo-de.
lafib¡a. -
Muy relacionadoscon la superficie de
Ias fibras de músculo esouelético se en-
cuentran núcleos más peqüeñosy más os-
curos, pertenecientesa las células satéli-
te, que son alargadasy aplanadasen di-
rección a la fibra muscular y están rodea-
das por la misma l¡ímina externa. Son
mioblastos persistentesy tienen impor-
tancia para la regeneración(véasecon ma-
yor detalle bajo histogénesisy regenera-
ción) y posiblemente también contribu-
yan al aumento de tamaño de un músculo
relacionadocon el entrenamiento.
Además de las miofibrillas. el sarco-
plasma contiene las organelase inclusio-
nes comunes. Cerca del polo nuclea¡ se
encuentran pequeños apa.ratosde Golgi y mitocondrias; estasúltimas también apa-
recen en hileras entre las miofibrillas. Me-
diante coloracionesespecialesse demues-
Fig.13-14.lmagende músculoesquelético tra la presenciade un fetículo sarcoplas-
captadaconmicroscopio electrónico.
Se obser- mático bien desarrollado que rodea las
va partede unafibramuscular y se identifican
confacilidad lasmiofibrillas,
ademásde las miofibrillas como una red y que cones-
bandasA, I, H, y la líneaZ. x18.000.(Cedido ponde al retículo endoplasmáticoliso en
porJ.V.Small.) otros tipos celulares. Por último, se en-
Miofibrillas
LíneaZ BandaA Banda I Banda H Línea M
308 TEJIDO MUSCULAR CAPíTU
Banda I posee una zona transversal menor, la
F.rt¿"_E {"1. ñe11,claro), glt qqtjlÉ¿tda
_b4lnqutee¡ los qoúes bistql-6giqqq,-y_ cd-,
detr_eqdqI cortada por q4a,lí4e_q_-Z!_te"!
definidalal. "s Zwischenscheibe, disco in-
termedio) o disco Z (dado que en tres di-
mensionesté-ve¡ue é'suna estructuracon
forma de disco), que s" tiñe con inténsi:
dad. En .elcentro.de la banda H se distin-
g-qeuna línea angosta,Ia línea M. EI seg-
'
mento ubicado entre dos líneas Z sucéili:
gitual déI sárcómero varfá con él estado
de contracción del músculo y alcanza
una extensión máxima de 3 pm, pero
disminuye hasta unos 1.,5pm con la con-
tracción. La lltngitud de Ls-hsndq A es-
constante, mientras quq..la banda I se ,
acorta durante Ia contracción. Por Io ge-
neral, con la microscopiaóptica i¿lo sán
-visibles las bandas A e I y,en o-c-asiqrte-q'
lqllnea Z (fí9. !.3-1.2).Todas las bandas
se distinguen con facilidad en Ias imáge-
Antiactinafluorescente nes captadascon microscopio electróni-
co (fig. L3-1,4).
Estudios estructurales y bioquímicos
Fig.13-15.Fotomicrografías de dos miofibri-
han dernostradoque la-miosina se locali=
llas aisladasde musculaturaesqueléticade
cone¡o,captadas pormicroscopiade con- za eatoda la banda A y la actina en la ban.
traste de fase (a) y de fluorescencia(b), res- -daI y en partes de la bandaA, Io cual tam-
pectivamente. Luegode seraisladas, lasmiofi- bién fue confirmado por métodos inmu-
brillasfueronsumergidas en unasolución de nohistoquímicos(fig. 13-15).
anticuerpo fluorescente de conejo.La
antiactina
antiactinase observafluorescente (blancoen la
imagen)en b y,comparado cona, se puedees-
tablecersu unióna lasbandasL Ambasimáge-
nesx2.000.(Cedidas porP Andersen y J.V.
Small.)
mún y en l¿ superficie citoplasmática in=
tern a se encuerrtra-Ja--proteína-distrofina
unida a las glucoproteínas de transmem-
cuentran cantidadesvariablesde glucóge- brana que fijan el sarcolemaa Ia matriz ex-
no y de gotasde Iípido dispersosen el ci- tracelular por anclaje con su dominio
toplasma. extracelular.A nivel aminoácidos,la dis-
Miofibrillas. El estriado- transve-rsal trofina se asemeja a Ia espectrina, que
carácteiGtiCldel músculo esquelé¡icq, confiere rigidez a la membranacelular de
que se distingue como bandas claras y los eritrocitos y es fundamental para el
ór"rrr"r a travZs de cada fibra, se debe al mantenimiento de la forma bicóncava
conieni¿o d-emio-fibrillas .estriadas, Las (véasecap. 10, pág. 239). Se cree que la
bandas transversalesse visualizan en la distrofina tiene un efecto similar sobre el
musculatura viva y en cortes coloreados sarcolema,como estabilizador mecánico,
(fig. 13-L2).Las bandas con mayor índi- y Ia carencia o modificación de la distro-
ce de refracción se tiñen con intensidad, fina posiblemente causen mayor predis-
mientras que las que poseen bajos índi- oosición a lesiones relacionadas con la
ces de refracción casi no se tiñen, Las óontracción,ademásde mayor permeabi-
bandas oscurasse denominan bandas A, lidad para los iones calcio. La distrofina
también es semejantea Ia alfaactinina y se
creeque ademástiene por función fijar los
filamentos de actina al sarcolema.En la
grave enfermedad distrofia muscular hay
carencia de distrofina o anomalías en su
luz polarizadaJfig.13-9d).Cadabanda A:. composición.
CAPITULO 13 TEJIDOMUSCULAR 309
Distrofia muscular
La distrofia muscular progresivainclu- hay disminución de la producción de dis-
ye un grupo de patologíasque se caracte- trofina o parte de ésta es anormal, pero
rizan por la degeneraciónprogresivade parcialmente fi.mcionante.Ambas oatolo-
Ias fibras muscularesesoueléticas.En la gías son heredita¡iasrecesivasligádasal
distrofia muscular de Duchenne los sín- sexo (la predisposición es portada por la
tomas se presentanva antesde los 5 años madre y la patologíase expresasólo en los
de edad, por ejempio porque el niño tie- hijos varones),pero alrededor de la tercera
ne dificultad para levantarsede la posi- parte de los casos se debe a mutaciones
ción de sentadoo acostadoy a partir de nuevas,sin presenciafamiliar previa de Ia
los 10-12añospor lo generalno puede ca- enfermedad.Se creeoue la mavor frecuen-
minar. La muerte suele ocurrir alrededor cia de mutaciones en el gen dé la distrofi-
de los 20 años. En la distrofia muscular na se debe a que hasta el momento es el
de Becker,más benigna,los síntomaspor gen más grande estudiado en el ser huma-
Io generalreciéncomienzanen los prime- no. Cabe destaca¡que existen otros tipos
ros años de Ia edad adulta v el oaciente de distrofia muscular, que no se deben a
sólo se ve lorzado a usar siila dó ruedas mutaciones del gen de la distrofina.
después de transcurridosunos 25 años. El conocimiento del gen de la distrofina
Además,Ia vida se puedeprolongarhasta y del producto genéücoabrela posibilidad
valoresnormales. del futu¡o tratamientopor tecnologíagené-
En la actualidadse sabeoue ta¡to la dis- tica de estasgravesenfermedades.Experi-
tofia de Duchennecomo lá de Beckerse mentos recientescon inserción de un gen
deben a mutaciones de los genesque codi- nonnal para dishofina (obtenido mediante
fican la distrofina. En la más grave,el üpo técnicasde recombinacióngenéüca)en fe-
de Duchenne,la mutaciónimplica que no tos murinos portadoresde la forma mutan-
se forma distrofina o que ésta es afuncio- te del gen ha¡r mostrado resultadospromi-
nante, mientras que en el tipo de Becker sorios, con curación de los ratones.
F i g .1 3 - 1 6 .l m á g e n e s d eml ú s c u l o e s q u e l é t i c o e n c o r t e s l o n g i t u d i n( a )l y t r a n s v e r s a(l b ) c a p -
tadascon microscopio electrónico. En a el cortees tangenciala una miofibrilla y se distinguenlos
elemenloslongitudinales del retículosarcoplasmático, ademásde las tríadastransversales. (Cedida
por P Flood) En b se observanlos elementosdel retículosarcoplasmático que rodeacada miofibri-
lla cortadaen sentidotransversal(Cedidapor J V Small)
iú¡rror
ri'T
',¡,:,
Q,
,{t1i ..'¡¡ L-
;*
ri:
¡& li
nt.id
Tríada
Retículosarcoolasmático
310 TEJIDO MUSCULAR CAPíTU
Además de las miofibrillas, sólo el re- (fig. 13-17). En los músculos de mamífe-
tículo sarcoplasmático presenta detalles ros hav 2 tríadas para cada sarcómero,
importantescon el microscopio electró- o,re rodeutr la mioiibrilla en la zorra co-
nico. Como se vio antes,correspondeal lresoondiente a la transición entre las
retículo endoplasmáticoliso de otras cé- bandas A e L
lulas, pero en la fibra muscular forma un Los túbulos T se comunican con el es-
reticulado denso de sarcotúbulos que ro- pacio extracelular, dado que son invagina-
dean cada miofibrilla ffigs. 13-16 y 13- ciones del sarcolema, lo cual se visualiza
17). Frentea las bandasA la mayor parte en las imágenes captadas con microscopio
de los sarcotúbulostranscurren en senti- electrónico.
do longitudinal y se anastomosanentre Mediante el túbulo un potencial de
sí, pero ademáspor fusión se forma un acción (una onda despolarizante) se
tubo más grande, el retículo de contacto. propaga con rapidez desde Ia superficie
Dos de estostubos rodeanla miofibrilla a de la fibra hasta el interior, donde favo-
ambos lados de un tubo más delgado,el rece Ia liberación de iones calcio desde
túbulo transversoo túbulo T. En coniun- el retículo hacia el sarcoplasma circun-
to, Ios tres tubos constituyenuna tríada dante.
Retículo
sarcoplasmático
Miofibrillas
BandaI
T
Túbulo
Retículo
BandaA sarcoplasmático
Retículode
contacto
TúbuloT
Microfibrillas
de colágeno
Fig. 13-17. Dibujoesque-
máticoque muestraen
tres dimensiones la ul-
traestructura de una fi-
bra muscular esqueléti-
ca. (SegúnKrstió.)
CAPITULO t3 TEJIDOMUSCULAR 311
Acoplamiento excitación-contracción en las fibras musculares esqueléticas:
base biomolecular
En correspondenciacon la trÍada hay bulos T se produce un cambio de la con-
una hendidura de sólo 15 nm entre las formación del receptor para DHP, lo que
membranasdel túbulo T y el retículo sar- a continuación causa Ia apertura de los
coplasmático.En la membranadel túbu- canalesde calcio en el recéptor de riano-
lo T se encuentra un complejo proteico dina de los pies de contacto,con el con-
compuestopor cuatro partículas.Se cree siguiente vaciamiento de iones calcio
que este complejo correspondeal recep- desde el retículo sarcoplasmático. La
tor de dihidroxipiridina (DHP), que fija función del receptor para DHP en la
DHP, un marcador de los canalesde cal- musculatura esqueléticase ha transfor-
cio dirigidos por potenciales.Las 4 partí- mado, desde ser un canal de calcio diri-
culas de la membrana del túbulo T (que gido por potenciales a ser un regulador
conforman el receptorpara DHP) se loca- dirigido por potenciales de los canales
lizan frente a las cuatro partículasprotei- de calcio del receptor de rianodina.
cas equivalentesen Ia membrana del re-
tículo de contacto (el retÍculo sarcoplas-
mático). Estasúltimas partículas forman
evaginacionesen el complejo de contac- Fig.13-18.Dibujoesquemático quemuestra
to, denominadas pies de contacto, dis- lospies de contactodel retículode contac-
puestos en hileras paralelasal eje longi- to y los receptoresDHPcontrapuestos en
el túbuloT (véaseel textoparalosdetalles).
tudinal del túbulo T (fig. 13-18).Las cua-
tro partículas de cada pie de contactore-
presentan el dominio citoplasmático de Tríada
cuatro conoles de liberación de calcio y
todo el complejo de estoscuatro canales
se denomina también receptor de riano-
dina (debido a que el complejo es capaz
de fijar especÍficamenteel alcaloide ve-
getal rianodina, que favorece Ia libera-
ción de calcio desde el retículo sarco- Retículo
de r'
plasmático). conlaclo
El retículo de contacto contiene en Ia
Iuz un material amorfo, compuestoen su
mayor parte por Ia proteína calsecuestri-
na que fija los iones calcio y así mantie-
ne la elevadaconcentraciónde éstos en
la luz reticular.
Pies de contacto
Se cree que cuando la onda de despo- que contienenel dihidroxipiridina
larización del potencial de acción ingre- receptorde (DHP)
sa en la fibra muscular a través de los tú- rianodina
Ei incremento de la concentraciónde mentos de dos tipos (figs. 13-9e-i y 13-19),
iones calcio en el sarcoplasmacausaen- con distintos espesor,longitud y composi-
tonces la contracción de la fibra muscular ción química. Los filamentos gruesos (de
(véasemás adelante).La denominación unos L4 nm) contienen miosina y se deno-
acoplamiento excitación-contracción in- minan filamentos de miosina. Los fila-
cluye estosprocesosdesde la excitación mentos /lnos (unos 7 nm) contienen actina
del sarcolema,con creacióndel potencial y se denominan filamentos de actina. En
d e a c c i ó ny s u d i f u s i ó n h a c i a e l i n t e r i o r Ia banda A los filamentos sruesos de mio-
de Lafibra muscular y la liberación de io- s i n a y J o s [ i l a m e n t o sf i n o s d e a c l i n a e s t á n
nes calcio, con Droducciónde la contrac- entremezclados (fig. 13-9e-i). Los filamen-
ción, que finaliza cuando la concentra- tos de miosina, de 1,5 ¡rm de largo, sólo
ción de iones calcio en el sarcoolasma llegan hasta el límite de las bandas A, don-
d i s m i n u y eh a s t aa l c a n z aer l n i v e l e n r e p o - de se afinan en los extremos. La distancia
so, por rebombeode los ionescalciohacia entre los filamentos de miosina es de unos
la luz del retículo de contacto mediante 45 nm. Los filamentos de actina oarten de
una Ca**-ATPasa. cada disco Z. desde donde se exlienden
Ultraestructura de las miofibrillas. Las L pm en cada dirección, para formar la
miofibrillas están formadas Dor miofila- banda I, pero continúan además dentro de
312 TEJIDOMUSCULAR CAPITU
Fig. 13-19. lmágenes, Sarcómero
captadascon microscopio
electrónico,de cortes
longitudinal y transver-
sal (a y b, respectivamen-
te) de músculo esquelé-
tico. Tantoen los cortes
longitudinales como en
los transversalesse Pue-
den diferenciarlos fila-
mentosgruesosde miosi-
na de los finos de actina.
En a se han marcadoto-
das las bandasy líneas
reconocibles.En b se ob-
serva un cofte transversal
a travésde las bandasA,
I y H, puestoque las 4
miofibrillascortadasa su
travésestán corridasa lo
largoentre sí en relación
con la localizaciónde los
miofilamentos.a x36.000.
b x50.000.(Ambasimá-
genes cedidaspor J.V.
Small.)
'ldtl
oarte de la banda A, donde se ubican entre miosina y de actina, mientras que en la
los filamentos de miosina. Faltan en la banda I sólo intervienen filamentos de ac-
banda H, compuestapor la porción media tina y en la bandaH, sólo los filamentosde
de la bandaA, hastadonde no lleganlos fi- miosina. La línea M correspondea una es-
lamentos de actina. En consecuencia, el tructura transversalde unión que une la
patrón de estrías transversalesse debe a Ia porción media, más gruesa,de los filamen-
superposición variable de Losdos tipos de tos de miosina. La línea M estácompuesta
filamentos. En la zona de superposiciónla por la proteÍnafijadora de miosina miome-
banda A se compone de filamentos de sina, a la que se une otra proteína fiiadora
CAPíTULO 1e TEJIDOMUSCULAR 313
Filamento
Filamentode de actina Porciónunida Po
actina a miosina elá
Filamen
-1oZ
Filamentode actina
Fig. 13-20. Dibujoesquemáticode la confor- sentanun citoesqueletorígido de la fibra.
mación propuesta para la línea Z en el mús- Los filamentos de estared forman una es- F i g . 1 3 - 2 1D
.
culo esquelético.(SegúnSchmalbruch y Rus- tructura anular alrededor del disco Z en máticode un r
ka, en Hirsch,Ruskay Sitte.) que muestral
cada miofibrilla, con la que se une. Estos
localizacióndr
anillos fibrosos también se unen median-
mentos de t¡t
te fibras transversalesentre las miofibri-
de miosina, denominada proteína C. En llas, por lo cual los discosZ y los sarcó-
Ios cortes transversales los filamentos for- meros mantienen su ubicación enf¡enta-
man un patrón hexagonal característico. da. Los filamentos intermedios estáncom-
En la zona de superposición cada filamen- puestospor desminaespecÍficade las cé-
to de miosina está rodeado por 6 filamen- lulas musculares(véasecap. 3, pág.98).
tos de actina (fig. 13-9i). Ademásde los filamentosde actinay de
Desde ios filamentos de miosina se ex- miosina, los sa¡cómeroscontienen un ter-
tiende en forma radial hacia los filamentos cer tipo de filamentos,designadosfilamen-
de actina una seúe de estructuras seme- tos de titina, compuestospor la proteínati-
jantes a puentes. Estas protrusiones faltan tina y de sólo 4 nm de espesor,por Io que
en una zona central de la banda H v esta no sevisualizanen Iasimágeneshabituales
región menos densa se denomina banda captadascon microscopioelectrónicoy de-
seudo H (en Ia figura 13-9e coinciden las ben ser demostradasmediantemétodoses-
bandas H y seudo Hl. Los puentes son de peciales. Se extienden desde Ia línea M
fundamental importancia para la interac- hastael disco Z. es decir a travésde Ia mi-
ción entre la actina y la miosina durante Ia tad del sarcómero.La titina es el polipépti-
contracción (véase más adelante). do de mayor tamaño conocido (de allí el
Se desconoce Ia estructura exacta de Ia nombretitina, del gr.titan, batalla,es decir,
Iínea Z. En los cortes longitudinales se ob- de la familia de guerrerostitanes) y cada
serva que cada filamento de actina se loca- molécula tiene una longitud equivalentea
liza frente al espacio intermedio entre 2 fi- la mitad del sarcómero.Un filamento de ti-
lamentos de actina sobre la cara oouesta de tina se fija al disco Z (f1g.13-21)y a un fi-
la lÍnea Z, y Ios filamentos de actiña opues- lamentode miosina.En consecuencia,el fi-
tos se relacionan a través de la línea Z me- lamento de titina tiene una Darteen la ban-
diante filamentos Z (fig. 13-9e). Se piensa da A, unida en toda su extensióncon un [i-
que cada filamento de actina forma la pun-
ta de una pirrímide, cuyas caras est¡ín for-
madas por los filamentos Z (fig. 13-20). Se
desconoce la comoosición de los filamen-
tos Z, pero gran parte del material electron-
denso del disco Z está compuesto por al-
faactinina, que une los filamentos de actina
con el disco Z por sus extremos positivos.
Los discos Z de las miofibrillas oeriféri-
cas están relacionados con estiucturas
ubicadas sobre la superficie interna del
sarcolema, que contiénen vinculina, ios
costámeros (laI. costa, costillas). Se for-
man así bandas costales o condensaciones
sobre la cara interna del sarcolema, con
una banda frente a cada disco Z en Ias
miofibrillas dyacentes. Los discos Z y, en Fig. 13-22. Di
consecuencia los sarcómeros, también se máticooue ml
mantienen en su lugar, uno frente al otro el sarcómero
a través de la fibra mediante una densa por deslizam
red de filamentos intermedios que repre- Contraído rante la contr
314 TEJIDO MUSCULAR CAPíTU
Fig. 13-23. Dibujoesque- de actina (fig. 13-23).Mediante el "método
máticode la configura- de las puntas de flecha" se demuestraque
ción molecular propues- los filamentos de actina poseenpolaridad,
ta para un filamento de
act¡na.
con un extremo positivo y un extremo ne-
gativo fvéasecap. 3, p. 91). El extremopo-
sitivo del filamento de actina se fija al dis-
co Z, mientras que el extremo negativo se
orienta en dirección opuesta.
A los filamentos de actina se fiian dos
lamento de miosina,y una porción elástica proteÍnasde importancia para Ia regula-
ubicada frente a Ia banda I entre los fila- ción de Ia contracción. La tropomiosina
mentosde actina.Por medios experimenta- es una molécula proteica de 40 nm (com-
les se ha demostradoque los filamentosde puestapor dos cadenaspolipeptídicasen-
titina mantienenlos fiiamentosde miosina roscadas)localizada en los surcos entre
en su lugar,en Ia parte media del sarcóme- dos cordones del filamento de actina F
ro. Ademásconfierenelasticidada las mio- (fig. 13-23).La troponina es una proteína
fibrillas y son la causade cierta resistencia globular grande localizada sobreun extre-
contra el estiramiento que presentan los mo de cadamoléculade tropomiosina,es
músculos, con independencia de Ia inte- decir, a intervalos de 40 nm sobre el fila-
racción entre actinay miosina. Se creeque mento de actina.La troponina se compo-
el objetivo perseguidoes contrarrestarla ne de tres polipéptidos: Ia troponina T se
sobreextensiónde las fibras musculares. une a la tropomiosina, a Ia que fiia así to-
Baseultraestructural de la contracción. do el compleio de la troponina; la troponi'
La longitud del sarcómerodisminuye du- na I se une a la actina,por lo que inhibe la
rante la contracción y mediante estudios unión de Ia actina con la miosina; la fro-
con difracción por rayos X y microscopia ponina C fila iones calcio.
electrónicase ha establecidoque los fila- Filamentos de miosina. Estánformados
mentos de actina se deslizanmós hacia el por Ia proteínamiosina, compuestospor
centro de la banda A durante el acorta- dos cadenaspesadasy cuatro cadenasli-
miento del sarcómero.De estemodo se ha- v i a n a s .L a m o l é c u l ad e m i o s i n ac o m p r e n -
cen más anBostaslas bandasH e I, mien- de una oorción con forma de bastóno "co-
tras oue la banda A mantieneinalteradasu 1a",en ia que las dos cadenaspesadas(ca-
longilud $ig. 13-22),Estemecanismode la da una es una hélice alfa) se enroscanpa-
contracción, denominado "hipótesis de ra formar una espiral arrollada que confie-
deslizamientode los filamentos" (ing. s1i- re a la cola ciertarigidez (fig. 13-2a).En el
ding filament hypothesis)tiene por resul- extremo de Ia cola las dos cadenaspesa-
tado total que los discosZ se acercanentre dasse seoaranv cadauna forma una óabe-
sí oor tracción v se acortatoda la miofibri- za de mi^osinaglobulu.. A cada cabezase
lla (por lo tantó, la fibra). adosandos cadenaslivianas, de las cuales
Base biomolecular de la contracción se piensa que la cadena liviana esencial
muscular.Los mecanismosmolecularesso- tiene importancia estructural para estabi-
bre los oue se basael deslizamientode los Iizar Ia cabezade miosina, mientras que la
filamentbs se han definido con claridad cadena liviana reguladora tiene impor-
mediante estudiosdetalladosde las carac- tancia para Ia regulaciónde la contracción
terísticasmolecularesde los miofilamentos. muscular en la musculatura 1iso,como ya
Filamentos de actina. En su mayor par- se vio, y en Ia musculaturaesqueléticater-
te estánformadospor una proteína globu- minó su evolución para "sólo" tener una
lar, la proteína G, que se polimeriza en dos función estabilizadora,similar a Ia de la
cordonesidénticos, Ia actina F (de fibras), c a d e n al i v i a n ae s e n c i a l .
que forman una espiral doble como dos ca- La forma descritade Ia miosina, con dos
denas de perlas. Esta espiral doble repre- cabezas,también se denomina miosina II,
senta la columna vertebral del filamento dado que existeuna forma con sólo una ca-
),"0",".
rolobulares
Fig. 13-24. Dibujoesque-
Porciónen forma de bastón,("cola")
f'
mát¡code la configura-
ción molecularde una
moléculade miosina. Moléculade miosina
CAPíTIJLO 13 TEJIDOMUSCULAR 315
beza,la miosina L La miosina I es Ia forma Fig. 13-25. D
máticodel em
oue interviene en la interacción entre acti-
) _.. miento de las
na y miosina en célu\asno musculares. I Fllamenlo de miosina el
Ante la acción débil de la enzima trin- oe mloslna
J mento de mic
s i n a s e d i v i d e n l a s m o l é c u l a sd e m i o s i ñ a
en dos subunidades, Ia meromiosina li-
viana [LMM) (ing. ]ight meromyosin) (gr.
meros, parte de), que representaIa mayor
parte de la porción con forma de bastón
de la molécula de miosina, v Ia meromio- Para formar el filamento de miosina se
sina pesada (HMM) (ing.-heory). com- empaquetan las moléculas de miosina con
puesta por el resto de Ia porción con for- las porciones con forma de bastón parale-
ma de bastónde la moléculade miosinay las al eje longitudinal del filamento. Las
las dos cabezasde miosina globulares(fig. cabezas de miosina se localizan en el ex-
L3-24).Ante Ia posterior acción de la en- tremo de la molécula orientada en direc-
zima papaína es posible escindir las cabe- ción opuesta a Ia línea M (fig. 13-9e y I) y
zas de la oorción con forma de bastón en con el microscopio electrónico se visuali-
la HMM, por lo que se han aislado las ca- zan como Ias mencionadas orotrusiones o
bezasy se han estudiadosus propiedades. puentes transversales.La polarización del
Cadauna de las cabezasde miosina ac- filamento de miosina, con las cabezas
túa como ATPasa.También ooseeun sitio orientadas en dirección oouesta a la línea
fiiador de actina. M es la causapor la cual-la parte cenl.ral
Filamentode actina
1 0n m
Fig. 13-26. Di
máticode la b
leculardel cic
puente transv
miosina (véas
oara los detall
316 TEJIDO MUSCULAR CAPITU
Ciclo del puente transversal de miosina: base biomolecular
Se describena continuación los pasos actina (fig. 1,3-26f).Una vez interrumpi-
del ciclo del puente transversal de mio- da la unión entre la cabezade miosina y
sina (fig. L3-26).En el músculo en repo- el filamento de actina se escinde ATP y
so (fig. 1.3-26a)la ATPasa de la cabeza la energía liberada vuelve a crear ten-
de miosina ha escindido el ATP (que en sión sobre la cabezade miosina al pasar
condiciones normales siempre se en- a la posición de 90., por Io que vueive al
cuentra en el músculo) y la energíalibe- punto de inicio del ciclo (fig. 1,3-26a),
rada oor la escisión causaun cambio de que se puede repetir, lo cual ocurre con
confolmación de la cabezade miosina, una velocidad de 2-3 vecespor segundo
oue en consecuencia"acumula tensión Dermanezcaactivada la fibra
mientras -oor
como u.r resorte" debido a haber varia- muscular mantenersela concentra-
do su dirección desde formar un ángulo ción elevada de iones calcio en el cito-
de unos 45ocon la porción con forma de sol. Así, la repetición del ciclo implica
bastón de la moléóula de miosina (con oue el filamento de actina es tracciona-
la cabezade miosina doblada en direc- do paso a paso (de 10 nm) hacia la ban-
ción de la banda H) hasta una posición da H. Cuando se interrumpe la activa-
con un ángulo de casi 90ocon el bastón ción disminuye la concentración de io-
de miosina. Los productos de la esci- nes calcio al nivel de reposo,donde los
sión de ATP, ADP y fosfato,permanecen iones de calcio ya no están unidos a la
unidos a la ATPasade la cabezade mio- troponina C. De este modo se modifica
sina, dado que ésta es incapaz de libe- Ia conformación del complejo de tropo-
rarlos antes de que Ia cabezade miosina nina hasta volver al estado de reposo,
se una a la actini. La activación de la fi- por Io que Ia molécula de tropomiosina
bra muscular por la producción de un otra vez cubre el sitio de unión para la
potencial de acción debido a un impul- cabezade miosina sobre el filamento de
so nervioso conduce, a través del aco- actina.De estemodo se detieneel ciclo
plamiento excitación-contracción (ya de la contracciónen el estadiode la fi-
descrito) a la liberación de iones calcio gura 1.3-26a, es decir se produce el rela-
desde el retículo sarcoplasmáticoy su jamiento del músculo. El ciclo de Ia
elevadaconcentraciónen el citosol. La contracción requiere, además de iones
unión de iones calcio a la troponina C calcio, la presenciade cantidadessufi-
desencadenaentoncesuna variación de cientes de ATP. El rigor mortis se debe
conformación del complejo de troponi- precisamentea que no se mantienen las
na, que desplaza Ia molécula de tropo- cantidadesnecesariasde ATP para man-
miosina de modo que ya no cubre el si- tener activa la Ca**-ATPasa(que dismi-
tio de unión para la miosina en la acti- nuye Ia concentración de iones calcio
na. Estoinicia el ciclo de Ia contracción, por bombeo hacia el interior del retícu-
dado que la cabezade miosina ahora se lo sarcoplasmático), por lo que las cabe-
fiia al sitio de unión libre sobre la acti- zas de miosina permanecenunidas a los
na (fig. 13-26b),lo que causaIa libera- filamentos de aitina, lo cual correspon-
ción de ADP y fosfato por la ATPasade de a Ia situaciónde Ia figura 13-26d,y el
la cabezade miosina (fig. 13-26c)y se- músculo no puede pasar al estadode re-
oara Ia cabeza de miosina de la tensa Iaiación.
-El
posición de 90., por lo que vuelve a su modelo descrito para Ia base bio-
posición de 45ocon el bastón de miosi- molecular del mecanismo de desliza-
na (fig. 13-26d).Como la cabezade mio- miento de los filamentos ha sido bien
sina estáunida a la actina durante el mo- estudiado experimentalmentey ha sido
vimiento, esto implica que el filamento aceptado en^ general, si bien existen
de actina es traccionado unos 10 nm en puntos de vista controvertidos respecto
dirección de la banda H (es obvio que a algunos de los detalles (p. ej., el grado
gran cantidad de cabezasde miosina de- de movilidad de las cabezasde miosina
sarrollan esta actividad al mismo tiem- y si parte de la porción con forma de
po). La inclinación de la cabezade mio- bastón también se desplaza).Es caracte-
sina hasta la posición de 45' implica rístico que los puentes transversalesde
también que los sitios de unión para las imágenes captadascon microscopio
ATP en la ATPasade la cabezade mio- electrónico se ubiquen en ángulos de
sina son accesibles,por lo que allí se fi- 45o o de 90o con los filamentos de acti-
ja ATP (fig. 13-26e),lo cual desencade- na, nunca en un ángulo intermedio, lo
na un cambio de conformación del sitio cual es una de las bases de la descrip-
de unión de la actina sobre Ia cabezade ción del modelo de movimiento de las
miosina y se interrumpe Ia unión con la cabezasde miosina.
CAPITULO to TEJIDO MUSCULAB 317
de Ia banda H carecede puentestransver- Fig. 13-27. Di
Fibra de músculoesouelético máticoque mr
s a l e sl,o q u e c r e aI a b a n d as e u d oH ( " z o n a
desnuda" de Huxlevl. una fibra nerl
ra por últimos
Las cabezasde miosina se disponen en muchasramif
espiral a lo largo del filamento de miosi- finas, cada ur
na, con una distancia de unos 40 nm entre cuales forma
cadagiro, dado que las moléculas de mio- motora termir
sina estándesplazadasen dirección longi- una fibra mus
tudinal y así se superponen(fig. 13-25). quelética.
Las moléculas de miosina son flexibles,
debido a la presenciade una "bisagra" en
Ia porción con forma de bastón, corres-
pondiente a la transición entre las mero-
Pfacamotoraterminal
miosinas liviana y pesada (fig. 13-26).
Fibra nerviosamotora
Además,la cabezade miosinaes móvil en
la transición hacia la porción con forma
de bastónde la moléculade miosina.
El deslizamiento de los filamentos de croscopia óptica que una fibra nerviosa
actina respectoa los filamentos de miosi- motora termina por dividirse en varias
na durante Ia contracción tiene luear de- cortas ramificacionesque establecencon-
b i d o a q u e l a m o l é c u l ad e m i o s i n ai e c o r r e tactos con las fibras musculares,a Ia ma-
el denominado ciclo del puente transver- nera de una mano con dedos irregulares,
sal de miosina (fig. 13-26),donde la cabe- con una ramificación para cada fibra mus-
za de miosina se fija a la actina,tracciona cuiar. La zona de contactocreaun pequeño
del filamentode actina fpor un cambio de engrosamientoen forma de placa,denomi-
la conformaciónde la mbléculade miosi- nado placa motora terminal (fig. 13-27),
na) y despuésinterrumpela unión. Al re- que también se puede demostrarmediante
petir esteciclo (unión, tracción,interrup- reacciónhistoquímica para Ia enzima ace-
ción de la unión) se desplaza("desliza") tilcolinesterasa,relacionada con la placa
el filamento de actina a lo largo del fila- terminal. Una fibra muscular poseesólo
mento de miosina en dirección hacia Ia una placa motora terminal localizadacer-
banda H, La energíadel procesoes produ- ca del centro lineal de Ia fibra. En Ia suoer-
cida por escisiónde ATP,catalizadapor la ficie de la fibra muscular se encuentran- ca-
porción ATPasade la cabezade miosina. vidades denominadas criptas sinápticas
primarias (fig. 13-28)donde se ubican las
Contacto neuromuscular terminalesaxónicas.En cada cripta sináp-
tica primaria también hay una serie de
La zona de contactoentre una fibra ner- hendiduras estrechasen Ia fibra muscular,
viosa motora y una fibra de músculo es- Ias criptas sinápticas secundarias.
quelético se denomina placa motora ter- Mediantemicroscopiaeiectrónicase vi-
minal. Por impregnacióncon salesmetáli- sualizan las células de Schwann (véase
cas se demuestraen preparadospara mi- con mayor detalle en el cap. 14) sobrela
Célulade Schwann---¿
Criptasinápticaprimaria
Criptassinápticassecundarias
@r.
Núcleode célula @
F i g . 1 3 - 2 8 .D i
máticodel asp
una placa mot
nal, visualizad
croscopio ele<
318 TEJIDO MUSCULAB CAPiTU
Miastenia gravis
En la enfermedad miastenia gravis las células T helper contribuyen como
(gr. mys, músculo; aesthenia, debili- en las reacciones inmunológicas nor-
dad) el paciente padece una definida m a l e s ( v é a s ec a p . 1 6 ) . E n e l t o - z o % d e
debilidad muscular con rápida transi- los oacientes se encuentra un tumor del
ción a impotencia. La enfermedad es timó, un timoma, y la timectomía es
autoinmuie, dado que se debe a la for- efectiva en algunos de los pacientes, so-
mación de anticuerpos contra el recep- bre todo en los jóvenes con evolución
tor de la acetilcolina en la placa motora reciente de la enfermedad. La patología
terminal, lo cual se demuestra en más reacciona bien con el tratamiento sinto-
del 90% de los pacientes con miastenia mático con inhibidores de la colineste-
generalizada. Esta enfermedad es Ia me- rasa (p. ej., piridostigmina), que dismi-
jor estudiada y mejor conocida de todas nuyen la degradación de acetilcolina y
las patologías inmunes. Los anticuer- así favorecen la estimulación de los re-
pos se fijan a los receptores y causan su ceptores accesibles para la acetilcolina
endocitosis (mediada por receptor, co- en la placa terminal. En los casos en
mo en la regulación hacia abaio común oue este tratamiento no es suficiente se
de los receptores) o bloquean los recep- inicia una terapéutica inmunosupreso-
tores (también pueden destruir los re- ra con hormonas corticosteroides o me-
ceptores por una acción mediada por el dicamentos citotóxicos. La sustancia ci-
complemento). Si bien la producción closporina, utilizada en casos severos,
de anticuerpos se debe a una respuesta actúá por inhibición de la síntesis y se-
inmune de linfocitos B, también los lin- creción de interleuquina 2 por Ios linfo-
focitos T intervienen activamente en el citos T helper por activación por los
desarrollo de la enfermedad, dado que linfocitos B.
s u p e r f i c i e d e l a l e r n l i n a l a x ó n i t : a( l i g s o r , L a a r ; e t i l t ; o l i r r ae s c l e g r a d a r i tar l c a b o
13-28) No se extienden hacia el interior c l e l r r - l o sp o r : o s t l i l i s t : g u n c l o s ¡ t o r l a e n z i -
d e l a s c r i p t a s s i n á p t i c a s .d o n d e e l a x o l e - m a a c e t i l c o l i n e s t e r a s al o c a l i z a t l ae u l a l á -
m a ( e s d e c i r , e l p l á s m a l e m a q u e r o c l e ae l n r i n a e x t e r n t ¿fri á n r i n a b a s a l ) d c l ¿ r m e m -
axón) v el sarcolema se encuentratt uno al b r a r r a l r o r s i r r á ¡ r lri rr I r r r i l t i u t - ri n t p r r l s o
lado d"elotro, sólo separados Por un esPa- n e r v i o s o l i b e r a l a r ; a n t i c . l a ed x a c ; t ad e a c e -
cio de unos 50 nm. La lámina externa clel t i l c o l i n a n e c e s a r i ap a r a p r o d u c i r r . r np o -
sarcolema recubre las criptas sináptir;as t e n c i a l d e a c ; r ; i ó ne t t e l s a r c o l e m a y u n a
primarias y secundarias. En el axoplasma única contract;ión de la fibra nerviosa.
se distineuen cúmulos de vesículas de La fibra nruscular )¡ los tendones po-
unos 50 ñm de diámetro, que correspon- seen órsallos terrninales sensitivos com-
den a las vesículas sinápticas de las si- plejos, denominados husos musculares y
naosis entre célulasnerviosas. t e n d i n o s o s ,q u e s e v e r á l t r : r ¡ nm a y o r d e l a -
La placa motora terminal se puede con- lle en el capítulo 14.
siderar una stnopsis modificada. Las vesí-
culas sinápticas contienen acetilcolina, Fibras musculares rojas, intermedias
o u e a c t ú a c o m o s u s t a n c i an e u r o l r a n s m i - y blancas
sora para la propagación del estímulo ner-
vioso desde el axón al sarcolema. El po- Los distintos mÍtsr;ulos esquelí:ticosva-
tencial de acción que llega a la placa ter- rían also en el color cuando se analizan en
minar libera acetiicolina de las vesículas frescoá simple vista. Además, las fibras
hacia la hendidura sináptica, donde las de un mismó mírsculo no tienen espesor
m o l é c u l a s d e a c e t i l c o l i n as e u n e n a r e c e p - uniforme. En los músculos rojos predomi-
tores para acetilcolina (de tipo nicotina) nan las fibras rojas, delgadas y de color
localizados sobre la membrana possináp- rojo oscuro, debido al gran contenido de
tica (sarcolema).La unión de la acetilcoli- mioglobina (el colorante rojo de los mús-
na con el receptor produce una aumento culos), En los músculos blancos predomi-
rápido y transitorio de la permeabilidad nan las fibras blancas, más gruesas y cla-
para los iones sodio, por lo que la mem- ras, debido al menor contenido de mioglo-
brana possináptica se despolariza como bina. Estos distintos tipos de fibras se de-
punto de inicio de un potencial de acción muestran y definen con facilidad median-
que se difunde por todo el sarcolema (véa- te métodos inmunohistoquímicos. La si-
s e c a p . 7 . p á g . 1 8 9 ) . E n c o n s e c u e n c i ae, l guiente descripción corresponde a las
receptor para acetilcolina es un canal ió- condiciones en el ser humano, donde casi
nico para sodio dirigido por un transmi- todos los músculos esqueléticos (al igual
CAPITULO 13 TEJIDOMUSCULAR 319
Fibrastipo I Fibrastipo llb Fibrastipo I
Fibrastipo lla Fibrastipo lla
Fig. 13-29. Fotomicrografías de músculo esquelético obtenidaspor biopsiadel músculovasto ex-
terno en una persona normal (a) y en un levantador de pesas (b). En amboscasos se realizóla
determinaciónhistoquímicade la ATPasamiofibrilara fin de diferenciarlos distintostipos de fibras
(la localizaciónenzimáticase observacomo un precipitadooscuro).Las fibrasdel tipo I son pobres
en ATPasa,mientrasque las fibrasde tipo lla y llb contienencantidadesvariablesde la enzima.Nó-
tese la gran hipertrofiaen el levantadorde pesas (b) de todos los tipos de fibras,en especiallas fi-
bras lla. (Cedidaspor H. Dahl.)
que todos los mamíferos)son del denomi- ta uno de los criterios más seguros,en hu-
nado "tipo twitch" (ing. estiramiento),es manos, para diferenciarlospor su ultraes-
decir, que reaccionan de acuerdo con Ia tructura de los otros dos tipos de fibras.
ley del todo o nada. Se entiende que und Las fibras tipo I son resistentesal agota-
fibra muscular siempre se contrae al mó- miento.
ximo ante el estímulo,si bien con diferen- Las fibras tipo IIa son un subgrupo de
te velocidad. Todaslas fibras musculares las fibras rápidas y contienen numerosas
pertenecientesa Ia misma unidad motora mitocondrias,si bien menos que las fi-
(son inervadas por el mismo axón) son de| bras tipo I. Este tipo de fibras también es
mismo tipo de fibra. resistente al agotamiento y parece ser
En la mayoría de los músculos esquelé- adecuadopara Ia actividad de resistencia
ticos humanos hay fibras twitch más rópi- y para contraccionesfuertes de escasadu-
dos y más lentas. Estos tipos de fibras se ración.
diferencian con mayor facilidad entre sí Las fibras tipo IIb se contraencon rapi-
mediante la determinación histoquímica dez y correspondena las fibras blancas.
de ATPasa miofibrilar (fig. ra-ZSj. Sobre Por lo generalson las fibras más gruesasy
esta base se clasifican las fibras en fibras forman grandes unidades motoras. Sólo
tipo I, pobres en ATPasa, y fibras tipo II, contienen escasasmitocondrias, respecto
ricas en ATPasa.Las fibras tipo II se clasi- a la masa miofibrilar. y se agotan muy
fican a su vez en dos subtipos; tipo IIa y pronto.
tipo IIb (un tercer tipo, IIc, posiblemente La mayor parte de los músculos se com-
sea un estadio previo de los otros dos ti- ponen de una mezcla de estos tres tipos
pos de fibras). de fibras, pero la distribución porcentual
Las fibras tipo I incluyen las fibras de los tres tipos es distinta de un múscu-
twitch lentas y correspondena las fibras lo a otro y de un individuo a otro. En las
ro.l'osmencionadas.A menudo son relati- contracciones musculares habituales se
vamente finas. y forman pequeñas unida- reclutan primero las fibras tipo I median-
des motoras. Las fibras tipo I poseen ele- te fuerzasde contracciónbajas,por ejem-
vado contenido de mitocondrias, en rela- plo las correspondientesa la actividad de
ción con las miofibrillas, Io cual represen- los músculos de posición erecta. Ante
320 TEJIDOMUSCULAR CAPíTU
fuerzas de contracción crecientes se in- forma de células satélite, a partir de las
corporan las fibras tipo II, pero la.cfil¡¿5 cuales se pueden desarrollar nuevas fibras
tipo IIb recién se incluyen ante aceleracio- durante la regeneración y quizá también
nes rápidas o pa-racontracciones máximas en relación con Ia hipertrofia de un mús-
de corta duración. culo debido a entrenamiento.
Se ha alcanzado mayor conocimiento
sobre la presencia de los tipos de fibras en Crecimiento y regeneración
seres humanos mediante aI estudio de
biopsias musculares. El diagnóstico defi- El aumento de la masa muscular que se
nitivo de las enfermedadesmuscul¿Iresse puede lograr con enuenamiento se debe
alcanza con el análisis de estas biopsias. fundamentalmente a aumento del espesor
La diferenciación entre los distintos tipos de las fibras musculares del músculo, al
de fibras tiene importancia fundamental igual que el crecimiento posnatal de los
en estoscasos. músculos durante el periodo de creci-
miento del individuo. En lo que respecta
Histogénesis aI aumento de espesor se debe a incre-
mento del espesorde cada una de las fi-
Toda la musculatura esquelética estria- bras musculares, dado que no se puede
da tiene origen mesodérmico y la mayoría demostrar aumento de Ia cantidad de fi-
se desarrolla a partir del mesodermo pa- bras musculares durante el crecimiento
raaxial, es decir, de los somitasdispuestos de los músculos despuésdel nacimiento.
en forma segmentaria. Sin embargo, la El crecimiento en espesor de cada fibra
musculatura de las fosas orbitarias y de Ia muscular se debe a aumento de la canti-
musculatura segmentariarelacionada con dad de miofibrillas. El crecimienro longi-
los arcos branquiales evolucionan in sifu tudinal de los músculos se debe a la pro-
del mesénquima. ducción de nuevos sarcómeros que se
Las primeras células que se diferencian agreganen la zona donde el músculo se
en dirécción de la musculatura esqueléti- une al tendón.
ca üenen forma de huso, los mioblastos, La cantidad de fibras en el músculo co-
con un único núcleo. No contienen miofi- mienza a disminuir a partir de los 25 años
brillas, pero sufren frecuente división' de edad y ademásse reduce el tamaño de
tras lo cual comienzan a fusion¿üsep¿Ira las fibras tipo II rápidas debido a atrofia.
forma¡ fibras musculares sinciüales mul- Ambos factores contribuyen a la menor
tinucladas, los miotubos, con núcleos fuerza muscular que se observa con la
centrales. Durante la diferenciación apa- edad.
recen zonas con estrías tra¡sversa-lesco- Regeneración. Después de la destruc-
mo consecuenciade la formación de mio- ción traumática de musculatura esquelé-
tica estriada, por ejemplo por "disten-
sión de fibras", comienza la regenera-
ción con la aparición de mioblastos,que
se presume se diferencian de las células
satélite. Los mioblastos se dividen en
forma activa y se fusionan para formar
tamaño de las fibras. Sin embargo, se cree nuevas fibras musculares del mismo mo-
que algunos mioblastos persisten bajo la do que durante la histogénesis' A conti-
nuación se producen miofibrillas estria-
das en las fibras recién formadas. En
Célulade adultos el poder de regeneración es lim-
músculocardíaco Discosintercalares
itado y en las lesiones importantes lasfi-
bras musculares lesionadasson reempla-
zadas por tejido conectivo. Despuésde
la pérdida se alcanza una compensación
poi hipertrofia de las fibras musculares
remanentes.
Músculo cardíaco
Fig. 13-30. Fotomicrogra-
fía de músculo caldíaco Las fibras musculares cardíacas están
coloreadocon azánPara compuestaspor célqlas que se ramifican y
poneren evidencia los
discos intercalares.Nó- forman en conjunto una red tridimensio-
teseel aspectosemeiante nal. Las células están unidas cola con co-
a unaescalera quePre- la mediante discos intercalares y el nú-
sentael discointercalar cleo tiene localización central (figs. 13-Ly
de la derecha.x440. 13-30),
CAPíTULO 13 TEJIDOMUSCULAR 321
F i g .1 3 - 3 1 .l m
músculo card
tada con micro
electrónico.Lo
traestructu rale
pondenfunda
te a los del mú
quelético Nóte
embargolas hi
mitocondrias (l
videnlos miof
en nacesque t
miofibrillas. x1Í
dida por Vodov
noyers.)
Características del músculo cardÍaco todo ei ancho de la fibra, pero suelen estar
con el microscopio óptico compuestos por varias pórciones longitu-
dinales desplazadas,por Io que presentan
Las fibras musculares cardíacas difie- tn aspecto semejante a una escalero. Los
ren de las esqueléticas en forma y tamaño. discos intercalares siempre se encuentran
En un corte transversal, su aspecto es me- a nivel de la parte ntedia de las bondos I.
nos regular, y en un corl.e l,ongitudinal. donde se Localizan las líneas Z. pero son
donde el recorrido de las fibras es bastan- más gruesas que éstas.
te paralelo, se observan ramificaciones
que se comunican con las fibras vecinas Ultraestructura de la musculatura
(fig. 13-1). EI diámetro de las fibras rara cardíaca
vez excede de 15 ¡rm. Los núcleos son
grandes, ovales y claros, y se encuentran La musculatura cardíaca tiene rasgos ul-
en la parte media de la célula (fig. 13-30). traestructurales fundamentales en común
Las células musculares cardíacas Doseen con la musculatura esquelética(fig. 13-31).
un sarcolema similar al de las fidras de La misma disposición precisa de los fila-
músculo esquelético, pero el sarcoplasma mentos de actina y de miosina crea el es-
es más abundante. Se distineue un nítido triado transversal. No obstante, ias miofi-
estriadolongitudinal con mióroscopioóp- brillas no presentan un empaquetamiento
tico. El patrón de estriado transversal y Ias tan denso como las fibras musculares es-
denominaciones de las distintas bandas queléticas, dado que hileras de mitocon-
corresponden a las de la musculatura es- drias y de elementos longitudinales del re-
quelética. Sin embargo, el estriado no es tícuio sarcoplasmático separan a los miofi-
tan notable. En cada polo nuclear se en- lamentos en haces paralelos anastomosa-
cuentra una pequeña zona de sarcoplasma dos (fig. 13-32). Las mitocondrias son mu-
con forma de clava, rica en mitocondrias, cho más numerosas y poseen gran canti-
que contiene un complejo de Golgi peque- dad de crestas(fig. 13-31). Además de ubi-
ño cerca del polo. En esta zona, y sobre to- c¿rrseen hileras entre las miofibrillas se al-
do en personas ancianas, se observan de- macenan en los polos nucleares. En los es-
pósitos de pigmento de iipofuscina. El pacios entre las mitocondrias se observan
sarcoplasma contiene más glucógeno que numerosas gotas de lípido y gránulos de
Ia musculatura esouelética. glucógeno, que son depósitos de energÍa.
Un rasgo característico es la presencia En el citoplasma de las células musculares
de los discos intercalares, ya nombrados, cardíacas de las aurículas, cerca de los po-
que se ven como gruesas líneas transver- los nucleares pueden aparecer vesÍculas
sales. Se distinguen mejor mediante tin- densas de unos 0,4 ¡rm de diámetro, los
ciones especiales (fig. 13-30) y a menudo gránulos auriculares, que contienen un
no se visualizan en preparados teñidos estadio previo de la hormona péptido na-
con HE. Los discos intercalares atraviesan triurético auricular (ANP) (ine. atrial na-
322 TEJIDO MUSCULAR CAPITU
triuretic peptid). Es un péptido secretado La contracción de la musculatura car-
oor las células musculares auriculares díaca tiene lugar con la misma forma de
óuandose estiran,por ejemplo cuando au- deslizamiento de filamenfos como en las
menta el volumen minuto cardíaco o Ia fibras de músculo esqueléticoy también se
oresión arterial.La hormona incrementala desencadenadebido a un aumento de la
óliminación de cloruro de sodio y aguapor concentación de iones calcio en las célu-
los riñones y tiene acción antihipertensiva las musculares cardíacas. Sin embargo,
fvéasecon mayor detalle en el cap. 15). existen ciertas diferenciasfuncionales re-
Los túbulos T tienen mayor diámetro lacionadas con el acoplamiento excita-
que en Ia musculatura esqueléticay siem- ción-contraccióny con la regulaciónde la
pre se localizan frente a las LíneasZ, por contracción.El potencial de acción difun-
lo que su número es menor que en la mus- de a travésdel sárcolemay se implanta por
culatura esquelética(fig. 13-32).Los túbu- medio de los túbulos T en la célula muscu-
los T de Ia musculatura cardíaca están lar cardíacaoara liberar iones calcio desde
unidos a intervalos por túbulos longitudi- el retículo salcoplasmático.Se creeque es-
nales con característicassimilares, pero ta contribución al aumento de Ia concen-
diámetro algo menor. Tienen la miima tración intracelular de iones calcio tiene
función que en Ia musculatura esqueléti- poca importancia para las fibras de la mus-
ca, es decir la propagación del potencial culatura cardíaca.Se cree que el mecanis-
de acción desde el sarcolemahacia el in- mo fundamentalse basaen que el poten-
terior de la fibra. cial de acción causaIa apertura de los ca-
EI retículo sarcoplasmático presenta nales iónicos de calcio dirigidos por po-
una conformación más simple, dado que tencialesen el sarcolema,tras lo cual la di-
está compuestopor un reticulado tubular fusión hacia el interior de los iones calcio
irregular que rodea los haces de miofila- incrementa Ia concentraciónen el citosol
mentos sin formar retículos de contacto v entonces oroduce Liberación de calcio
interrelacionados (cisternas terminales). inducida oir calcio desde el retículo sar-
En los túbulos T sólo se forman pequeñas coplasmático (de modo similar al de las
exoansionesaisladasen los extremosde células musculareslisasl. La unión de los
la ied tubular, sin creaciónde trÍadasdife- iones calcio a la troponina C causala con-
renciadas.En estaszonasel retículo sar- siguientecontracción.Sin embargo,Ia ca-
coolasmáticoestablececontactoscon los pacidadde las célulasmuscularesparaal-
túbulosT similaresa los de Ia musculatu- macena-riones calcio en el retículo sarco-
ra esouelética.Parte del retículo endo- plasmáticoes mucho menor que en las fi-
plasmáticoliso, el retículo sarcoplasmáti- bras musculares esqueléticas debido al
co corbular (fr. corbeille,canasto),se loca- menor tamaño del reiículo, por Io que Ia
Iiza justo por debajo del sarcolemay for- difusión de iones calcio desde el espacio
ma con él contactossemeiantes. extracelularhacia el interior de Ia célula
es importante para alcanzar una concen-
tración de iones calcio de nivel suficiente
Fig.13-32.Dibujo esquemáticoquemuestra en el citosol para desencadenaruna con-
en tresdimensionesla conformaciónultraes- tracción de clerta intensidad en Ia célula
tructuralde unafibra muscularcardíaca. muscular. En realidad, durante la activi-
(SegúnFawcett y McNutt) dad en reposodel corazónsólo se produce
unión de iones calcio a Ia troponina C en
Mitocondria
Retículosarcoolasmático TúbuloT
Túbulo
T cantidad eouivalentea alrededorde la mi-
tad de la áctivación máxima del aparato
contráctilde la célula muscularcardíaca,
por lo que hay capacidadpara incrementar
la fuerza de la contraccióna travésdel au-
mento de la concentraciónintracelular de
calcio. Esta forma de graduar la fuerza de
contracción difiere de la musculatura es-
ouelética. en la cual una fibra muscular
slemore se contrae al m¡áximocuando se
despólarizapero se pueden obtenerdiver-
sosgradosde contracciónen el músculo al
variar la cantidad de fibras reclutadas y al
permitir que se contraigan en asincronía.
Estaforma de graduaciónno es posible en
el músculo cardíaco,donde toda la masa
muscular de la aurícula o del ventrículo se
activa en forma casi simultánea.
La finalización de Ia contracción li.ene
Retículosarcoplasmático lugar por disminución de la concentra-
CAPiTULO IJ TEJIDO MUSCULAR 323
Ultraestructura de los discos intercala-
res. En los cortes longitudinales se visua-
lizan los extremosopuestosde las células
musculares cardíacas interdigitados (fig.
13-33).Las membranascelularesadyacen-
tes están separadaspor un espacio inter-
celular de unos 20 nm y presentan, en
consecuencia, un transcurso alternante
transversal y longitudinal (fig. 13-34).
Frente a las porciones transversalesse
distingue un material electrondensoen el
citoplasmasubsarcolémicoal cual se fijan
los filamentos de actina de los sarcómeros
adyacentes,que correspondea la localiza-
ción del disco intercalar frente al disco Z.
Los filamentos intermedios también se fi-
Fig. l3-33. Dibuloesquemáticodel aspectotri- ian a estascondensacionescon caracterís-
dimensionalde un disco intercalar en el mús- ticas de adhesiones focales. Además se
culo cardíaco. (SegúnPochey Lindner.) observandesmosomastÍpicos y por fin al-
gunos nexos aisladospequeñosa la altura
de las partestransversalesy más nexos de
mayor tamaño a la altura de la sección
ción de iones calcio en el citosol. Dor re- Iongitudinal de los discos intercalares,
captación al retículo sarcoplasmático donde las céluias vecinas se encuentran
mediado por una ATPasa activada por lado a lado. Frente a los nexos falta el ma-
calcio o por extracción por bombeo me- terial electrondensoen el citoplasma, da-
diante la corresoondiente bomba en el do que los nexosrepresentanzonasdonde
sarcolema. Además se extraen iones cal- el potencial de acción es transmitido rápi-
cio de la célula por medio de un meca- damentede una célula a otra por difusión
nismo de intercambio sodio-potasio, por de iones a travésdel canal de cadauno de
el cual tres iones de sodio del espacio ex- Ios conexonesdel nexo (véasecap. 6, pág.
tracelular se intercambian con un ion 1,67).De este modo se obtiene un acopla-
calcio del citosol (la energía es provista miento eléctrico de toda Ia masamuscular
por el gradiente de sodio a t¡avés del sar- cardíaca,que así funcionacomo un sinci-
colema y, en consecuencia depende de la tio. Los desmosomasy las adhesionesfo-
bomba Na*-K*). Los iones calcio introdu- cales tiene por función unir entre sÍ por
cidos por bombeo en el retículo sarco- anclaje las miofibrillas de fibras adyac-en-
plasmático se unen a la calsecuestrina, tes, para que Ia intensidad de la contrac-
de modo similar a la musculatura esque- ción se transmita de una célula a otra.
lética. Dado oue el corazón se conirae Sistema de transmisión de impulsos
gran cantidad de veces por minuto y que del corazón. Si bien un potencial de ac-
oor Io tanto cada célula muscular cardía- ción se ouede difundir a toda la muscula-
óa se despolariza con igual frecuencia, tura caidÍaca por medio de los nexos.
debido a la aoertura de los canales de existe ademásun sistemade células mus-
calcio dirigidos por potenciales en el sar- culares cardíacasmodificadas,capacesde
colema habrá un inereso bastante cons- difundir potencialesde acción con mayor
tante de iones calcioá la célula, y en con-
secuencia también estarán activos cons-
tantemente los mecanismos de extrac-
ción por bombeo, Io cual genera una per- 19
manente graduación del depósito de cal-
cio en el retículo sarcoolasmático. El in-
cremenlo de la frecuenóia cardÍaca causa
una concentración de iones calcio relati-
va mavor en el citosol. debido al aumen- I
to del-flujo hacia el interior desde el es-
pacio extracelular por unidad de tiempo,
en consecuencia aumenta el deoósito de
iones calcio en el retÍculo sarcoolasmáti-
co durante la fase de relaiación. De este Fig. 13-30. lm
modo aumenta la cantidad de iones cal- disco intercal
4
cio que se liberan en la siguiente despo- t músculo card
larización y también la intensidad de Ia tada con micrc
contracción en cada célula muscular car- electrónico.(C
díaca, y de todo el músculo cardíaco. E. Holm Niels
324 TEJIDO MUSCULAR CAPITU
rapidez que las fibras comunes. Este siste- Crecimiento y regeneración
ma de transmisión de impulsos permite
que ia contracción de las aurículas y de Después del parto, el corazón sólo crece
los ventrículos tenga lugar en la secuencia l:or aumento de tamaño de cada célula
mejor dirigida para la función de bombeo. muscular cardíaca, dado que como se
El sistema de transmisión de impulsos y mencionó antes no hay mitosis en la vida
las células musculares especializadas que extrauterina. En el adulto las células al-
intervienen en él se verán con mayor de- canzan un diámetro de unos 15 ¡rm, pero
talle en el caoítulo 15. donde se describe en condiciones patológicas (o en casos de
el corazón como órgano y donde además enlrenamientomuy intensivo y prolonga-
se analizarán su irrigación e inervación. do) con mayor sobrecarga cardíaca se pro-
duce hipertrofia del corazón con aumento
Histogénesis de la masa muscular. Esta tiene lugar por
incremento del espesor de las fibras hasta
La musculatura cardíaca evoluciona a casi 20 ¡lm y también por aumento de Ia
oartir de mioblastos oue difunden desde la lonsitud de cada fibra. El corazón carece
porción del mesodermo esplácnico que ro- de óapacidad regenerativa debido la falta
dea los tubos cardíacos limitados Dor en- de caoacidad mitótica en las células mus-
dotelio. Las célulascontinúan su diuisión cularés cardíacas después del nacimiento
por mitosis durante y después de finaliza- y a que no existen células satélite, como en
da la diferenciación hasta poco antes del la muscuiatura esouelética.En consecuen-
nacimiento, lo cual indica condiciones cia, en casos de isquemia prolongada, por
bien distintas de las oue rodean el desarro- ejemplo, por oclusión coronaria, se produ-
Ilo de Ia musculatuia esquelética. en la ce necrosis tisular, donde el defecto en la
c u a l l a s c é l u l a s y a d i f e r e n ó i a d a sy a n o s e musculatura cardíaca es reemplazado por
dividen. ni siouiera durante la vida fetal. una cicatriz de teiido conectivo.
Cuestionario sobre tejido muscular
1. ¿Cómose denomina¡ los tres tipos esqueléticay qué enfermedadgrave
de músculosv en oué se diferencian produce su carenciao Ia variación
respectoa la iocali2aciónde los nú- de su composición?
cleos y al estriado transversal? 11. ¿Quécomponentesconformanuna
2. ¿Quétipo de contactocelula¡ une a tríada en Ia musculatu¡a esquelética?
Ias célulasmusculareslisas entre sí? 12. ¿Quéfunción cumplen las tríadasen
3. ¿Cómose organizael citoesqueletode la musculaturaesouelética?
Ias células musculareslisas (filamen- 13. ¿Cómose organizanlos filamentos
tos intermedios,condensaciones cito- de actina y de miosina en un sarcó-
plasmáticasy placasde inserción)? mero?
4. Intente describir el mecanismo de 14. Describael mecanismo de desliza-
contracciónde las célulasmuscula- miento de filamentos para la con-
res lisas. tracción de la musculatura esquelé-
5. ¿Cómodesencadena la enzima miosi- tica.
na-cadenalivianaouinasa la contrac- 15. ¿Cómopueden las cabezasde miosi-
ción en la célula muscular lisa? na "traccionar" Ios filamentos de ac-
6. ¿Quéimportancia tienen los nexos tina hacia la zona H?
que unen las células muscularesli- L6. Describa el aspectoultraestructural
sas? de una placa motora terminal.
7. ¿Cómose denomina la unidad es- L7. ¿Córnose definen los tres tipos de fi-
tructural y funcional de la muscula- bras de la musculatura esqueléticay
tura esqueléticay cómo se define? cuáles son sus característicasfunda-
B.¿Québandaso líneastransversalesse mentales?
distinguen en un buen preparado pa- 18. ¿Cuáles el aspectode los discosin-
ra microscopia óptica de una fibra tercalaresen los prcparados para mi-
muscular esquelética? croscopia óptica de musculatura es-
9. ¿Quévariaciones presentan las longi- queléticay dónde se localizanres-
tudes de las bandasA e I durante la pecto del estriado transversal?
contracción? 19. ¿Quéimportancia tienen los nexos
10. ¿Cómose denomina la proteínarela- en la musculatura cardÍaca?
cionada con la superficie citoplasmá- 20. ¿Cuáles la capacidad regenerativa
tica del sarcolemaen la musculatura del corazón despuésdel nacimiento?
CAPITULO 13 TEJIDO MUSCULAR 325
326 TEJIDOMUSCULAR CAPíTU
Potrebbero piacerti anche
- Biologia 1NES - DocenteDocumento26 pagineBiologia 1NES - DocentePaulaMed280% (5)
- Biologia 1NES - DocenteDocumento26 pagineBiologia 1NES - DocentePaulaMed280% (5)
- Cejas y OccifrontDocumento2 pagineCejas y OccifrontCinthy GC100% (2)
- 01 Tejido MuscularDocumento28 pagine01 Tejido Muscularmanuela castroNessuna valutazione finora
- Sistema MusculoesqueleticoDocumento39 pagineSistema MusculoesqueleticoMARIA FERNANDA OLIVEROS ANDRADE100% (1)
- Músculo EsqueléticoDocumento23 pagineMúsculo EsqueléticoLeslie LópezNessuna valutazione finora
- Infografía Biología CelularDocumento1 paginaInfografía Biología CelularabrilataolivaresNessuna valutazione finora
- Explicación Músculo Liso-2 PDFDocumento8 pagineExplicación Músculo Liso-2 PDFMaysa FreitasNessuna valutazione finora
- 2023 RUSTEIN BIO 13 TejidoMuscularDocumento5 pagine2023 RUSTEIN BIO 13 TejidoMuscularzambonjuliangabrielNessuna valutazione finora
- Anexo 2. Taller de SIstemas BiomecánicosDocumento24 pagineAnexo 2. Taller de SIstemas BiomecánicosMayra Alejandra Quintero ValenciaNessuna valutazione finora
- Tejido Muscular (Sixto Javier Reto Zapata) PDFDocumento1 paginaTejido Muscular (Sixto Javier Reto Zapata) PDFSixto ZapataNessuna valutazione finora
- Ilovepdf MergedDocumento63 pagineIlovepdf MergedCamila DominguezNessuna valutazione finora
- Generalidades Del Sistema MuscularDocumento19 pagineGeneralidades Del Sistema MuscularMARTINEZ SANCHEZ JULIAN ELIAS100% (1)
- Tema 3 - Sistema MuscularDocumento19 pagineTema 3 - Sistema MuscularluciaNessuna valutazione finora
- Tejido MuscularDocumento21 pagineTejido MuscularJaribel ToribioNessuna valutazione finora
- Anatomofisiologia Aplicada Clase 8Documento31 pagineAnatomofisiologia Aplicada Clase 8scarletNessuna valutazione finora
- Práctica. Tejido MuscularDocumento3 paginePráctica. Tejido MuscularEliot Ismael AcostaNessuna valutazione finora
- Musculo 1Documento24 pagineMusculo 1Rafael PazNessuna valutazione finora
- Tema 19 (Tejido Muscular)Documento37 pagineTema 19 (Tejido Muscular)Carlos Daniel Cárdenas SantosNessuna valutazione finora
- Trabajo de LaboratorioDocumento3 pagineTrabajo de Laboratoriopia veronNessuna valutazione finora
- Mapa Conceptual-Sistema MuscularDocumento4 pagineMapa Conceptual-Sistema MuscularlicmiriampaxtianumNessuna valutazione finora
- EXPOSICIÓNDocumento28 pagineEXPOSICIÓNMILENI NATALIN VILLANUEVA BARRANTESNessuna valutazione finora
- Tejido MuscularDocumento11 pagineTejido MuscularGraciela Romero PalacioNessuna valutazione finora
- Lostejidos PDFDocumento3 pagineLostejidos PDFIcdj MontalvoNessuna valutazione finora
- Cuestionario MúsculoDocumento8 pagineCuestionario MúsculoDARWIN RUDY CAMPERO GUZMÁNNessuna valutazione finora
- Tejido MuscularDocumento13 pagineTejido MusculardanielaNessuna valutazione finora
- Sistema MuscularDocumento3 pagineSistema MuscularMeytiNessuna valutazione finora
- Clase 13 TejidoMuscularDocumento30 pagineClase 13 TejidoMuscularSolange Colina0% (1)
- Sistema Muscular-Victoria SanchezDocumento4 pagineSistema Muscular-Victoria SanchezVictoria Sanchez NietoNessuna valutazione finora
- Anatomia Miologia 2021Documento9 pagineAnatomia Miologia 2021IzeckNessuna valutazione finora
- Tarea 2. Soporte y MovimientoDocumento50 pagineTarea 2. Soporte y Movimientojhon sanchez100% (1)
- Tejido MuscularDocumento63 pagineTejido MuscularDannoxNessuna valutazione finora
- Musculos 1Documento5 pagineMusculos 1Ees 5 Esteban EcheverriaNessuna valutazione finora
- ANATOMIA Apunte OAM - MUSCULOS - 2023Documento5 pagineANATOMIA Apunte OAM - MUSCULOS - 2023yesscanteroeventosNessuna valutazione finora
- Tejido MuscularDocumento21 pagineTejido MuscularGonzález Albarrán AlejandraNessuna valutazione finora
- Taller Teorico de Sistema MuscularDocumento13 pagineTaller Teorico de Sistema MuscularMarianella HerreraNessuna valutazione finora
- HistoDocumento10 pagineHistomeilyarteaga8Nessuna valutazione finora
- Guía Sistema MuscularDocumento15 pagineGuía Sistema MuscularCHELCY DIAZ GARCESNessuna valutazione finora
- Erika Cachupud Morfofisiologia - DeberesDocumento13 pagineErika Cachupud Morfofisiologia - DeberesErika CachupudNessuna valutazione finora
- Musculo CardiacoDocumento2 pagineMusculo CardiacoFrancesca PupoNessuna valutazione finora
- Anatomia de La Piel PaulinaDocumento2 pagineAnatomia de La Piel PaulinaROCIO MADRIGAL HNessuna valutazione finora
- Tejido Muscular (Documento29 pagineTejido Muscular (naomi orozcoNessuna valutazione finora
- Sistema MuscularDocumento5 pagineSistema MuscularPauloMazziniNessuna valutazione finora
- Cudro DiferencialDocumento6 pagineCudro Diferencialbeltran romeroNessuna valutazione finora
- Funcion de Los Musculos SuperficialesDocumento56 pagineFuncion de Los Musculos SuperficialesMaria Acosta100% (3)
- Tejido MuscularDocumento20 pagineTejido MuscularSebastián MendezNessuna valutazione finora
- Tipos de TejidosDocumento2 pagineTipos de Tejidosreyes122515Nessuna valutazione finora
- Tejido Muscular 198668 Downloable 1121332Documento5 pagineTejido Muscular 198668 Downloable 1121332Nayire Barboza HuatangareNessuna valutazione finora
- Anatomia de Los SistemasDocumento50 pagineAnatomia de Los SistemasP f bNessuna valutazione finora
- Sistema Musculo Esqueletico LISTODocumento5 pagineSistema Musculo Esqueletico LISTOODALIS JANETH PACHECO RIVERANessuna valutazione finora
- Valoracion Anatomia Tema 1Documento8 pagineValoracion Anatomia Tema 1verde04 04Nessuna valutazione finora
- Cuadro Comparativo Entre Los Sistemas Oseo y MuscularDocumento4 pagineCuadro Comparativo Entre Los Sistemas Oseo y MuscularDahiana Borja100% (3)
- Tejido MuscularDocumento12 pagineTejido MuscularJoselin America Pérez PolancoNessuna valutazione finora
- Sistema Muscular Bases y Fisiología MuscularDocumento10 pagineSistema Muscular Bases y Fisiología MuscularManuel Gonzales BriosNessuna valutazione finora
- Aparato MusculoesqueleticoDocumento2 pagineAparato MusculoesqueleticoisamarNessuna valutazione finora
- Mapa MasajeDocumento3 pagineMapa MasajeWalter DizuelaNessuna valutazione finora
- Seminario 10 - Músculo Liso y EsqueléticoDocumento17 pagineSeminario 10 - Músculo Liso y EsqueléticoAdriano MagallanesNessuna valutazione finora
- Histo UpaoDocumento1 paginaHisto UpaoFernando RodriguezNessuna valutazione finora
- Introducción A La HistologíaDocumento11 pagineIntroducción A La HistologíaRomiglio AlejandroNessuna valutazione finora
- Cuadro de TejidosDocumento2 pagineCuadro de Tejidosestefania figueroaNessuna valutazione finora
- 2) Músculo (Stuart Fox 2011)Documento44 pagine2) Músculo (Stuart Fox 2011)Guillermo AmengualNessuna valutazione finora
- Tejido Muscular ResumenDocumento21 pagineTejido Muscular ResumenEl KurkoNessuna valutazione finora
- Cuadro ComparativoDocumento10 pagineCuadro ComparativoNicolas GuzmanNessuna valutazione finora
- Anatomía artística 5: Articulaciones y funciones muscularesDa EverandAnatomía artística 5: Articulaciones y funciones muscularesValutazione: 4 su 5 stelle4/5 (1)
- 02.CAPÍTULO - 7-Glándulas y Secreción PDFDocumento20 pagine02.CAPÍTULO - 7-Glándulas y Secreción PDFLauraSeoaneIvanacNessuna valutazione finora
- Ciencias Naturales y TICDocumento382 pagineCiencias Naturales y TICJorge Alejandro Léon Sanchez100% (1)
- 01.CAPÍTULO - 6-Epitelio PDFDocumento20 pagine01.CAPÍTULO - 6-Epitelio PDFLauraSeoaneIvanacNessuna valutazione finora
- Ciencias Naturales y TICDocumento382 pagineCiencias Naturales y TICJorge Alejandro Léon Sanchez100% (1)
- CAPÍTULO - 13-Tejido Muscular PDFDocumento28 pagineCAPÍTULO - 13-Tejido Muscular PDFLauraSeoaneIvanac100% (1)
- 02.CAPÍTULO - 7-Glándulas y Secreción PDFDocumento20 pagine02.CAPÍTULO - 7-Glándulas y Secreción PDFLauraSeoaneIvanacNessuna valutazione finora
- Clasificacion Cinco ReinosDocumento21 pagineClasificacion Cinco ReinosLauraSeoaneIvanacNessuna valutazione finora
- 01.CAPÍTULO - 6-Epitelio PDFDocumento20 pagine01.CAPÍTULO - 6-Epitelio PDFLauraSeoaneIvanacNessuna valutazione finora
- Órgano Genital FemeninoDocumento11 pagineÓrgano Genital FemeninoLauraSeoaneIvanacNessuna valutazione finora
- Tejidos Vasculares PrimariosDocumento1 paginaTejidos Vasculares PrimariosLauraSeoaneIvanacNessuna valutazione finora
- Homeostasis y Retroalimentacion l419032013 - 0000Documento1 paginaHomeostasis y Retroalimentacion l419032013 - 0000LauraSeoaneIvanacNessuna valutazione finora
- Biologia I PDFDocumento37 pagineBiologia I PDFSusy SegoviaNessuna valutazione finora
- PDFDocumento4 paginePDFLauraSeoaneIvanacNessuna valutazione finora
- Comunicación NeuronalDocumento15 pagineComunicación NeuronalFlavio Daniel Morales HernandezNessuna valutazione finora
- La Vía de Síntesis de Los Ácidos Grasos Ocurre en El CitoplasmaDocumento7 pagineLa Vía de Síntesis de Los Ácidos Grasos Ocurre en El CitoplasmaLauraSeoaneIvanacNessuna valutazione finora
- Control Biológico de Enfermedades de Plantas en América Latina y El CaribeDocumento404 pagineControl Biológico de Enfermedades de Plantas en América Latina y El CaribeLauraSeoaneIvanacNessuna valutazione finora
- NCh1596-1999 Cortes Menores o BásicosDocumento21 pagineNCh1596-1999 Cortes Menores o BásicosGianluca CattaneoNessuna valutazione finora
- Ciclo Celular. HistoloiaDocumento11 pagineCiclo Celular. Histoloiarafa favelaNessuna valutazione finora
- Sistema MuscularDocumento24 pagineSistema MuscularVladimir MQNessuna valutazione finora
- Protocolo de Evaluación en Parálisis FacialDocumento7 pagineProtocolo de Evaluación en Parálisis FacialCarla BelénNessuna valutazione finora
- Músculo EsternocleidomastoideoDocumento6 pagineMúsculo EsternocleidomastoideoAngela Mejia ArmasNessuna valutazione finora
- Banco de Preguntas Anatomía IDocumento4 pagineBanco de Preguntas Anatomía IKevin AgNessuna valutazione finora
- Músculos Del AntebrazoDocumento3 pagineMúsculos Del AntebrazoFer BrionesNessuna valutazione finora
- SuprahioideoDocumento2 pagineSuprahioideoGissette Tatiana Bautista Gomez100% (1)
- La Contracción MuscularDocumento28 pagineLa Contracción Muscularirly1935100% (1)
- Tabla Reflejos NeurológicosDocumento2 pagineTabla Reflejos NeurológicosMiriam LujanNessuna valutazione finora
- Movimientos Articulares y Músculos ImplicadosDocumento21 pagineMovimientos Articulares y Músculos ImplicadosJacob Gonzalez Barrios50% (6)
- Antebrazo Muñeca y Mano 6Documento56 pagineAntebrazo Muñeca y Mano 6api-370208767% (3)
- Cingulo Toracico y HombroDocumento51 pagineCingulo Toracico y HombroKatherine LavieraNessuna valutazione finora
- 06 AntebrazoDocumento60 pagine06 AntebrazoLizy FloresNessuna valutazione finora
- Musculos AnatomíaDocumento2 pagineMusculos AnatomíaLucia GarayNessuna valutazione finora
- Musculos Escapulohumerales.Documento2 pagineMusculos Escapulohumerales.Manuel DavidNessuna valutazione finora
- Examen TESDocumento1 paginaExamen TESMelanie GonzalezNessuna valutazione finora
- 03 Evaluacion de Postura-1Documento7 pagine03 Evaluacion de Postura-1Isaac Alegria100% (1)
- Tipos de Contracciones Musculares PDFDocumento2 pagineTipos de Contracciones Musculares PDFSara RiosNessuna valutazione finora
- Anatomia Miemb. Inferior CedevDocumento12 pagineAnatomia Miemb. Inferior CedevFranco AlonsoNessuna valutazione finora
- El Tono MuscularDocumento2 pagineEl Tono MuscularjhensssNessuna valutazione finora
- 13-Musculos de La Cadera PDFDocumento3 pagine13-Musculos de La Cadera PDFSofi Campos100% (1)
- Anatomía Del NadadorDocumento30 pagineAnatomía Del Nadadorjuan guerra100% (1)
- Conceptos CoreDocumento31 pagineConceptos CoreSrta. IntrusaNessuna valutazione finora
- MusculosDocumento29 pagineMusculosVanesa RamirezNessuna valutazione finora
- Nervios IntercostalesDocumento5 pagineNervios Intercostalesluis angelNessuna valutazione finora
- Cintura PélvicaDocumento71 pagineCintura Pélvicagio_pedroniNessuna valutazione finora
- DermatomaDocumento3 pagineDermatomabetofisicosudNessuna valutazione finora