Potrebbero piacerti anche
- Laporan Aktualisasi: by Raymond Mangapultua Janius JansenDocumento12 pagineLaporan Aktualisasi: by Raymond Mangapultua Janius JansenClarita BangunNessuna valutazione finora
- Vibrio Vulnificus: SymptomsDocumento3 pagineVibrio Vulnificus: SymptomsClarita BangunNessuna valutazione finora
- Definisi InfrastrukturDocumento7 pagineDefinisi InfrastrukturClarita BangunNessuna valutazione finora
- Desain Perhitungan IpalDocumento2 pagineDesain Perhitungan IpalClarita BangunNessuna valutazione finora
- Proposal and Marketing Plan: Subtitle Text HereDocumento2 pagineProposal and Marketing Plan: Subtitle Text HereClarita BangunNessuna valutazione finora
- The Germ Theory of DiseaseDocumento5 pagineThe Germ Theory of DiseaseClarita BangunNessuna valutazione finora
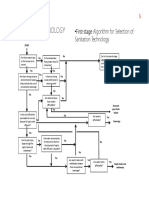
- FlowChart SeleksiTeknologiSANITASI PDFDocumento3 pagineFlowChart SeleksiTeknologiSANITASI PDFClarita BangunNessuna valutazione finora
- 08 Gaya GayaDocumento22 pagine08 Gaya GayaClarita BangunNessuna valutazione finora
- John Snow (1813-1858) and CholeraDocumento3 pagineJohn Snow (1813-1858) and CholeraClarita BangunNessuna valutazione finora
- KSLING1Documento5 pagineKSLING1Clarita BangunNessuna valutazione finora
- Grafik X (Jarak) Terhadap Y (Kedalaman)Documento18 pagineGrafik X (Jarak) Terhadap Y (Kedalaman)Clarita BangunNessuna valutazione finora
- What's Protectionism?: OutlineDocumento4 pagineWhat's Protectionism?: OutlineClarita BangunNessuna valutazione finora
- English ConjunctionDocumento8 pagineEnglish ConjunctionClarita BangunNessuna valutazione finora
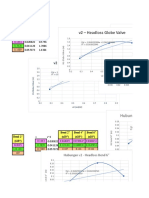
- v2 - Headloss Gate ValveDocumento8 paginev2 - Headloss Gate ValveClarita BangunNessuna valutazione finora
- NAME: Clarita MONTH: November: SundayDocumento2 pagineNAME: Clarita MONTH: November: SundayClarita BangunNessuna valutazione finora
- Presentation SkillDocumento107 paginePresentation SkillClarita BangunNessuna valutazione finora
- Academic WritingDocumento88 pagineAcademic WritingClarita BangunNessuna valutazione finora
- Arca Majora 2 LicenseDocumento1 paginaArca Majora 2 LicenseClarita BangunNessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5795)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Gasem MH HANTA Salatiga 2019Documento43 pagineGasem MH HANTA Salatiga 2019ChristianNessuna valutazione finora
- Rat and Mice Infestation Reporting in Ohio - Google SearchDocumento1 paginaRat and Mice Infestation Reporting in Ohio - Google SearchmakeupbydesignNessuna valutazione finora
- Viral Infections of The Gastrointestinal Tract and Viral Infections of The Genitourinary SystemDocumento14 pagineViral Infections of The Gastrointestinal Tract and Viral Infections of The Genitourinary SystemDARLENE CLAIRE ANDEZANessuna valutazione finora
- Nbme 16 AnsDocumento17 pagineNbme 16 Ansnithin shenoi0% (1)
- Viral Infection AHEADDocumento24 pagineViral Infection AHEADShiena Mae PelagioNessuna valutazione finora
- Hantavirus Epi AlertDocumento4 pagineHantavirus Epi AlertSutirtho MukherjiNessuna valutazione finora
- The History of Biological Warfare 1Documento4 pagineThe History of Biological Warfare 1Thonieroce Apryle Jey MorelosNessuna valutazione finora
- Principles of Molecular VirologyDocumento8 paginePrinciples of Molecular VirologyAJCannNessuna valutazione finora
- Case 12-2020: A 24-Year-Old Man With Fever, Cough, and DyspneaDocumento10 pagineCase 12-2020: A 24-Year-Old Man With Fever, Cough, and DyspneatabareeNessuna valutazione finora
- Similarity Report IoT 30AUG22Documento18 pagineSimilarity Report IoT 30AUG22Ajay NainNessuna valutazione finora
- Microbiology Assignment. ADocumento26 pagineMicrobiology Assignment. ACynthia AbbangNessuna valutazione finora
- 2017 - Gastrointestinal Tract As Entry Route For Hantavirus Infection - FrontiersDocumento9 pagine2017 - Gastrointestinal Tract As Entry Route For Hantavirus Infection - FrontiersFlorin BranisteNessuna valutazione finora
- Viral Hemorrhagic Fever Viral Hemorrhagic FeverDocumento44 pagineViral Hemorrhagic Fever Viral Hemorrhagic FeverBlessing Paul100% (1)
- Emerging and Reemerging Infectious DiseasesDocumento13 pagineEmerging and Reemerging Infectious DiseasesSayu100% (1)
- Emerging and Reemerging InfectionsDocumento57 pagineEmerging and Reemerging Infectionstummalapalli venkateswara raoNessuna valutazione finora
- Bioterrorism Agents/Diseases: Student Name: Ayesha AndleebDocumento15 pagineBioterrorism Agents/Diseases: Student Name: Ayesha AndleebAisha rana100% (1)
- Case Study 1.1. The Nurse Who Told A Lie: Did The Nurse Do The Right Thing? Was The Doctor's Advice Good Advice?Documento12 pagineCase Study 1.1. The Nurse Who Told A Lie: Did The Nurse Do The Right Thing? Was The Doctor's Advice Good Advice?Payal NaiduNessuna valutazione finora
- Microbiology Practice Test IIIDocumento14 pagineMicrobiology Practice Test IIISidharta ChatterjeeNessuna valutazione finora
- A Topic Presentation ON: Emerging Communicable DiseasesDocumento52 pagineA Topic Presentation ON: Emerging Communicable DiseasesGiri SivaNessuna valutazione finora
- Hantavirus Disease: ImportanceDocumento11 pagineHantavirus Disease: Importancemanju laNessuna valutazione finora
- BunyavirusDocumento24 pagineBunyavirusManny GabrielNessuna valutazione finora
- Hemorrhagic FeverDocumento3 pagineHemorrhagic FeverDenis QosjaNessuna valutazione finora
- Hanta Vi Ru S Inf Ection SDocumento20 pagineHanta Vi Ru S Inf Ection Stummalapalli venkateswara rao100% (3)
- Arduino Guided Smart Rodent Trap (Trapuino)Documento9 pagineArduino Guided Smart Rodent Trap (Trapuino)Eunice Nicole Masecampo LautingcoNessuna valutazione finora
- Viral Infections of HumansDocumento89 pagineViral Infections of HumansMark Vincent JanoyogNessuna valutazione finora
- Spillover: Animal Infections and The Next Human Pandemic by David QuammenDocumento5 pagineSpillover: Animal Infections and The Next Human Pandemic by David QuammenSouthern California Public Radio40% (5)
- Important Lists of Helminths: Non-Bile Stained Eggs Operculated Eggs Don't Float in Sat. Salt SolutionDocumento10 pagineImportant Lists of Helminths: Non-Bile Stained Eggs Operculated Eggs Don't Float in Sat. Salt SolutionAjay ShresthaNessuna valutazione finora