Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocumento5 pagineAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Abbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFDocumento2 pagineAbbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFflashjetNessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Abbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFDocumento5 pagineAbbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFflashjetNessuna valutazione finora
- Nampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFDocumento4 pagineNampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFflashjetNessuna valutazione finora
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
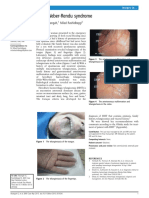
- Abangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeDocumento2 pagineAbangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeflashjetNessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Nascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestDocumento4 pagineNascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestflashjetNessuna valutazione finora
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Nana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFDocumento2 pagineNana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFflashjetNessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocumento4 pagineAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetNessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Nascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntDocumento3 pagineNascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntflashjetNessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocumento5 pagineAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetNessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Namburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFDocumento4 pagineNamburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFflashjetNessuna valutazione finora
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Nathwani - 2013 - A Complicated Case of AnticoagulationDocumento2 pagineNathwani - 2013 - A Complicated Case of AnticoagulationflashjetNessuna valutazione finora
- Nascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesDocumento3 pagineNascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesflashjetNessuna valutazione finora
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocumento4 pagineAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetNessuna valutazione finora
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Narula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFDocumento3 pagineNarula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFflashjetNessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Nasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeDocumento3 pagineNasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeflashjetNessuna valutazione finora
- Nascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseDocumento5 pagineNascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseflashjetNessuna valutazione finora
- Stroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437Documento29 pagineStroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437MaryamNessuna valutazione finora
- Nasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildDocumento3 pagineNasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildflashjetNessuna valutazione finora
- Nassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctDocumento3 pagineNassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctflashjetNessuna valutazione finora
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Idiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewDocumento7 pagineIdiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewflashjetNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- Naveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarDocumento4 pagineNaveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarflashjetNessuna valutazione finora
- Natrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaDocumento6 pagineNatrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaflashjetNessuna valutazione finora
- Nassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirDocumento3 pagineNassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirflashjetNessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
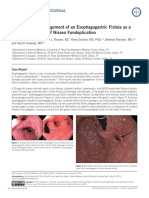
- Naveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationDocumento2 pagineNaveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationflashjetNessuna valutazione finora
- Nguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2Documento36 pagineNguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2flashjetNessuna valutazione finora
- Notes 200430 195226 0c2 PDFDocumento19 pagineNotes 200430 195226 0c2 PDFflashjetNessuna valutazione finora
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (120)
- Notes 190915 213814 Ea7 PDFDocumento1 paginaNotes 190915 213814 Ea7 PDFflashjetNessuna valutazione finora
- Naveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisDocumento4 pagineNaveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisflashjetNessuna valutazione finora
- Nguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisDocumento4 pagineNguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisflashjetNessuna valutazione finora
- Pensions Reforms at NSSFDocumento4 paginePensions Reforms at NSSFcliffmogere2348Nessuna valutazione finora
- Appendix VIII - Civil and Structural Scope of WorkDocumento140 pagineAppendix VIII - Civil and Structural Scope of WorkjaganrajNessuna valutazione finora
- Tutorial Sheet 4Documento2 pagineTutorial Sheet 4Syed YousufuddinNessuna valutazione finora
- Plane Bearings: Material Indicator Shape or SeriesDocumento4 paginePlane Bearings: Material Indicator Shape or SeriesJuan LoaizaNessuna valutazione finora
- Chery Amulet 1,6 Engine Service ManualDocumento76 pagineChery Amulet 1,6 Engine Service ManualG x HxhNessuna valutazione finora
- Literature Review On Procrastination Impact On Academic PerformanceDocumento2 pagineLiterature Review On Procrastination Impact On Academic Performancemomi100% (1)
- Modified Pfirrmann Grading System For Lumbar Intervertebral Disc DegenerationDocumento6 pagineModified Pfirrmann Grading System For Lumbar Intervertebral Disc DegenerationZack StylesNessuna valutazione finora
- Automated Hematology Cell Counters 12-09-2023Documento161 pagineAutomated Hematology Cell Counters 12-09-2023Tom JohnathanNessuna valutazione finora
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Mitsubishi Galant 4g63 Engine Repair ManualDocumento29 pagineMitsubishi Galant 4g63 Engine Repair ManualMoaed Kanbar100% (1)
- April 12th Test PDFDocumento32 pagineApril 12th Test PDFهخه •Nessuna valutazione finora
- Past PapaersDocumento515 paginePast PapaersShariq ShaNessuna valutazione finora
- Hyundai forklift specification dimensionsDocumento6 pagineHyundai forklift specification dimensionsАлександр ОлейникNessuna valutazione finora
- Indian Pharmacopiea 2007Documento73 pagineIndian Pharmacopiea 2007Deepak ShahNessuna valutazione finora
- Apcosil 605Documento1 paginaApcosil 605rihtak123Nessuna valutazione finora
- Healthy Voice: by Dan VascDocumento22 pagineHealthy Voice: by Dan VascscoutjohnyNessuna valutazione finora
- Johnson Et Al.2006. Felidae PhylogenyDocumento6 pagineJohnson Et Al.2006. Felidae Phylogenyaspergillus_jallNessuna valutazione finora
- LV CB Catalogue PDFDocumento260 pagineLV CB Catalogue PDFBoobesh Kumar SekarNessuna valutazione finora
- IRIS 2017 DOG Treatment Recommendations 09may18Documento15 pagineIRIS 2017 DOG Treatment Recommendations 09may18crilala23Nessuna valutazione finora
- UM16000 Flame Detector Installation Guide EnglishDocumento12 pagineUM16000 Flame Detector Installation Guide Englishmohamed.rescoNessuna valutazione finora
- By Kerri Rivera, Kimberly Mcdaniel and Daniel Bender: Healing The Symptoms Known As Autism Second EditionDocumento24 pagineBy Kerri Rivera, Kimberly Mcdaniel and Daniel Bender: Healing The Symptoms Known As Autism Second EditionMarilen De Leon EbradaNessuna valutazione finora
- Ats, TT, Tig: Anti Tetanus Serum, Tetanus Toxoid, Tetanus Immuno-GlobulinDocumento15 pagineAts, TT, Tig: Anti Tetanus Serum, Tetanus Toxoid, Tetanus Immuno-Globulinjisoo100% (2)
- Nihms 223615 PDFDocumento15 pagineNihms 223615 PDFElay PedrosoNessuna valutazione finora
- Berca Engineering SUBMITTEDDocumento20 pagineBerca Engineering SUBMITTEDJohandika FerbiantokoNessuna valutazione finora
- Earth First! Climbers Guild Vol. 2Documento16 pagineEarth First! Climbers Guild Vol. 2CascadiaDefenders100% (2)
- Tap ChangerDocumento5 pagineTap ChangerRavindra AngalNessuna valutazione finora
- Child Rearing Practices Articles India & ChinaDocumento14 pagineChild Rearing Practices Articles India & ChinaGeorgiana GattinaNessuna valutazione finora
- Emerging Foodborne Enteric Bacterial Pathogens: A ReviewDocumento11 pagineEmerging Foodborne Enteric Bacterial Pathogens: A ReviewAngelica Alejandra De La Torre AnayaNessuna valutazione finora
- Implementation of Swachh Bharat in MysoreDocumento13 pagineImplementation of Swachh Bharat in MysoreDipyaman ChoudhuryNessuna valutazione finora
- Spares Parts Cannibalisation ProcedureDocumento3 pagineSpares Parts Cannibalisation ProcedureThang Nguyen Hung100% (1)
- Filipino Nurses United OrientationDocumento30 pagineFilipino Nurses United OrientationErlinda PosadasNessuna valutazione finora
- Tales from Both Sides of the Brain: A Life in NeuroscienceDa EverandTales from Both Sides of the Brain: A Life in NeuroscienceValutazione: 3 su 5 stelle3/5 (18)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDa EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisValutazione: 3.5 su 5 stelle3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDa EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityValutazione: 4 su 5 stelle4/5 (3)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDa EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesValutazione: 4.5 su 5 stelle4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDa EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsValutazione: 4.5 su 5 stelle4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDa EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceValutazione: 4.5 su 5 stelle4.5/5 (515)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDa Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessValutazione: 4 su 5 stelle4/5 (33)