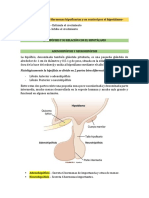
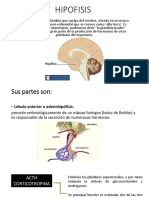
Potrebbero piacerti anche
- La Infertilidad No Es Sólo Un Problema De La ParejaDa EverandLa Infertilidad No Es Sólo Un Problema De La ParejaNessuna valutazione finora
- Hormonas de La Fisiología Reproductiva en BovinosDocumento13 pagineHormonas de La Fisiología Reproductiva en BovinosJesus Gomez Cortizzo100% (1)
- HORMONAS Del HipotalamoDocumento7 pagineHORMONAS Del HipotalamoLevi Hernandez LopezNessuna valutazione finora
- Membrana CelularDocumento28 pagineMembrana CelularAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Hemorragia ObstetricaDocumento24 pagineHemorragia ObstetricaDennis CRamNessuna valutazione finora
- Unidad Feto Placentaria.Documento33 pagineUnidad Feto Placentaria.JEAG1196% (23)
- Curso Liturgia para JovenesDocumento23 pagineCurso Liturgia para JovenesAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Diapositivas HORMONASDocumento170 pagineDiapositivas HORMONASmary_pacasNessuna valutazione finora
- Resumen Cap. 76 - Hormonas Hipofisarias y Su Control Por El HipotálamoDocumento11 pagineResumen Cap. 76 - Hormonas Hipofisarias y Su Control Por El Hipotálamojohan manuel candelario de la rosaNessuna valutazione finora
- Desprendimiento de Placenta.Documento9 pagineDesprendimiento de Placenta.Marlen ValdezNessuna valutazione finora
- Hormonas HipotalámicasDocumento7 pagineHormonas HipotalámicasJisselle Then AlvarezNessuna valutazione finora
- REPRODUCCIONDocumento20 pagineREPRODUCCIONKatherine WilsonNessuna valutazione finora
- Hormonas de La PlacentaDocumento9 pagineHormonas de La PlacentaLisseth MosqueraNessuna valutazione finora
- Sistema EndocrinoDocumento8 pagineSistema EndocrinoJosé Luis Gonzáles SosaNessuna valutazione finora
- Hormonas PlacentariasDocumento6 pagineHormonas PlacentariasangelfountainNessuna valutazione finora
- HipofisisDocumento14 pagineHipofisisyoseannysgomez1617Nessuna valutazione finora
- Anatomia HipotalamoDocumento3 pagineAnatomia HipotalamoGerardo Reyes BonifantNessuna valutazione finora
- Funciones Que Controlan Las HormonasDocumento6 pagineFunciones Que Controlan Las HormonasCami JuliánNessuna valutazione finora
- Sistema Endocrino ResumenDocumento12 pagineSistema Endocrino ResumenJuliett OsorioNessuna valutazione finora
- Hormonas Durante El EmbarazoDocumento10 pagineHormonas Durante El Embarazotrebin_16Nessuna valutazione finora
- Unidad Feto PlacentariaDocumento33 pagineUnidad Feto PlacentariaPaulo Linares PercaNessuna valutazione finora
- Prolactina Jade IbáñezDocumento14 pagineProlactina Jade IbáñezsandiaNessuna valutazione finora
- Sistema NeuroendocrinoDocumento6 pagineSistema NeuroendocrinoTināNessuna valutazione finora
- Resumen Del Eje Hipotalamo-HipofisisDocumento5 pagineResumen Del Eje Hipotalamo-HipofisisanahyrasconNessuna valutazione finora
- Repaso ParcialDocumento7 pagineRepaso ParcialHoracio PosdeleyNessuna valutazione finora
- Endocrinología Reproductiva de La HembraDocumento4 pagineEndocrinología Reproductiva de La HembraKaren SalazarNessuna valutazione finora
- T4 FubicoDocumento9 pagineT4 FubicoBelén Alatriste FortesNessuna valutazione finora
- Fisiología Reproductiva de La YeguaDocumento34 pagineFisiología Reproductiva de La YeguaHarold Christopher Cortez CercadoNessuna valutazione finora
- Cuestionario N11Documento2 pagineCuestionario N11Ronaldo Cuarez OrtizNessuna valutazione finora
- APUNTES Endocr Del Climaterio y MenopausiaDocumento13 pagineAPUNTES Endocr Del Climaterio y MenopausiaANA ELIZABETH MENDO URBINANessuna valutazione finora
- Fisiologia 1Documento5 pagineFisiologia 1mariela caisana riberaNessuna valutazione finora
- Hormonas Hipofisiarias y Su Control Por HipotalamoDocumento13 pagineHormonas Hipofisiarias y Su Control Por HipotalamoCodfishxx76Nessuna valutazione finora
- HipotalamoDocumento11 pagineHipotalamoBen CórdovaNessuna valutazione finora
- 3 Resumen Hormonas HipofisariasDocumento9 pagine3 Resumen Hormonas HipofisariasCelesteAngelicaCoelloPorrasNessuna valutazione finora
- Hipotalamo HipofisisDocumento19 pagineHipotalamo Hipofisislaura fonsecaNessuna valutazione finora
- Resumen Sistema EndocrinoDocumento7 pagineResumen Sistema EndocrinoMafe TrianaNessuna valutazione finora
- Hormonas PlacentariasDocumento10 pagineHormonas PlacentariasJosé Manuel Torres AvendañoNessuna valutazione finora
- 6.hormonas Placentarias - Ginecologia IDocumento42 pagine6.hormonas Placentarias - Ginecologia IIvan Emanuel Caballero PazNessuna valutazione finora
- Endocrinologia ReproductivaDocumento78 pagineEndocrinologia ReproductivaMartha Rueda MoncadaNessuna valutazione finora
- Comunicación EndócrinaDocumento6 pagineComunicación EndócrinaJose Pelaez RosasNessuna valutazione finora
- Test Sobre HormonasDocumento3 pagineTest Sobre Hormonasjclavijo6311Nessuna valutazione finora
- Hormonas Placentarias (Documento7 pagineHormonas Placentarias (claudia elena escobar contrerasNessuna valutazione finora
- Fsiologia Del Aparato Reproductor Femenino Grupo MELISSADocumento6 pagineFsiologia Del Aparato Reproductor Femenino Grupo MELISSACristhel Rios arnezNessuna valutazione finora
- Eje Hipotálamo HipófisisDocumento33 pagineEje Hipotálamo HipófisisABRAHAMJONessuna valutazione finora
- Fisiopato ResumenDocumento4 pagineFisiopato ResumenLizeth Yucra ArapaNessuna valutazione finora
- 3.3anatomía Del Eje Hipotálamo-Hipófisis. TiroidesDocumento6 pagine3.3anatomía Del Eje Hipotálamo-Hipófisis. TiroidesCristhel RiosNessuna valutazione finora
- EXPO Endocrinologia FetalDocumento65 pagineEXPO Endocrinologia FetalGrecia Torres ValderramaNessuna valutazione finora
- Endocrinologia en La ReproducciónDocumento30 pagineEndocrinologia en La ReproducciónChihiro JanNessuna valutazione finora
- Hormonas y GlandulaDocumento6 pagineHormonas y GlandulaBRIGITTE ESTEFANIA DELGADO GUILCAPINessuna valutazione finora
- FX Endocrina de La Placenta.Documento22 pagineFX Endocrina de La Placenta.api-3720763100% (3)
- Endocrinología de La GestaciónDocumento4 pagineEndocrinología de La GestaciónHollow Sv0% (1)
- Síndrome de Secreción Inadecuada ADHDocumento37 pagineSíndrome de Secreción Inadecuada ADHErick BetetaNessuna valutazione finora
- Hormonas Que Sintetizan en La AdenohipofisisDocumento34 pagineHormonas Que Sintetizan en La AdenohipofisisTomás Emmanuel Hernández CuevasNessuna valutazione finora
- SSyR - FSHDocumento50 pagineSSyR - FSHLu FinochiettoNessuna valutazione finora
- Semiología Eje Hipófisis - Hipotálamo, Tiroides.Documento73 pagineSemiología Eje Hipófisis - Hipotálamo, Tiroides.gabrielasaenz931Nessuna valutazione finora
- Sistema Regulador Hipotálamo-HipófisisDocumento19 pagineSistema Regulador Hipotálamo-HipófisisVeronica CandoNessuna valutazione finora
- Gine Clase 1Documento7 pagineGine Clase 1Claudio MuñozNessuna valutazione finora
- Endocrinologia ReproductivaDocumento55 pagineEndocrinologia ReproductivaGraciela NuñeNessuna valutazione finora
- RESUMEN de HormonasDocumento4 pagineRESUMEN de HormonasYezseniaaAC'Nessuna valutazione finora
- ANATOMOFISIOLOGIADocumento9 pagineANATOMOFISIOLOGIAJohan Coello RomeroNessuna valutazione finora
- Compendio Fisio 2pDocumento87 pagineCompendio Fisio 2pSarah CalderonNessuna valutazione finora
- HipofisisDocumento5 pagineHipofisisAna Vidal O.Nessuna valutazione finora
- HIPOFISISDocumento10 pagineHIPOFISISFrancis NicoleNessuna valutazione finora
- ExtraDocumento8 pagineExtrakiara CabreraNessuna valutazione finora
- 3 - Aparato Reproductor Hembra y MachoDocumento79 pagine3 - Aparato Reproductor Hembra y MachoAracely SanchezNessuna valutazione finora
- Metabolismo y Respiración Celular. NúcleoDocumento39 pagineMetabolismo y Respiración Celular. NúcleoAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Manual de Terminologia MédicaDocumento132 pagineManual de Terminologia MédicaAdriana Arianny Diaz Castañeda100% (2)
- 4 Esquema 2017 4to AñoDocumento15 pagine4 Esquema 2017 4to AñoAdriana Arianny Diaz CastañedaNessuna valutazione finora
- REENCUENTRO ANATÓMICO (Sistema Respiratorio)Documento18 pagineREENCUENTRO ANATÓMICO (Sistema Respiratorio)Adriana Arianny Diaz CastañedaNessuna valutazione finora
- P1 Farmacologia IIDocumento20 pagineP1 Farmacologia IIAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Estructura de La BibliaDocumento2 pagineEstructura de La BibliaAdriana Arianny Diaz CastañedaNessuna valutazione finora
- P1. MFH IV. Curso 2014 PDFDocumento14 pagineP1. MFH IV. Curso 2014 PDFAlejandro PadillaNessuna valutazione finora
- Farmacologia GeneralDocumento215 pagineFarmacologia GeneralAdriana Arianny Diaz Castañeda100% (1)
- 2 Do Año Esquema Docente 2019Documento12 pagine2 Do Año Esquema Docente 2019Adriana Arianny Diaz CastañedaNessuna valutazione finora
- Glandulas SuprarrenalesDocumento31 pagineGlandulas SuprarrenalesAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Glandulas SuprarrenalesDocumento17 pagineGlandulas SuprarrenalesAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Procederes Básicos de EnfermeríaDocumento44 pagineProcederes Básicos de EnfermeríaAdriana Arianny Diaz Castañeda100% (1)
- Flujograma de Atencion 1 PDFDocumento1 paginaFlujograma de Atencion 1 PDFAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Desarrollo EmbriofetalDocumento14 pagineDesarrollo EmbriofetalAdriana Arianny Diaz Castañeda100% (1)
- Sistema EndocrinoDocumento22 pagineSistema EndocrinoAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Desarrollo Pre EmbrionarioDocumento21 pagineDesarrollo Pre EmbrionarioAdriana Arianny Diaz CastañedaNessuna valutazione finora
- Rciu LibroDocumento36 pagineRciu LibroYolpa Figueroa Porras100% (1)
- Parto EutocicoDocumento63 pagineParto EutocicoDr. Jaime R. Castro AguirreNessuna valutazione finora
- Práctica 2. Disco Trilaminar y Periodo EmbrionarioDocumento11 paginePráctica 2. Disco Trilaminar y Periodo EmbrionarioJhon Miguel Barragan CascoNessuna valutazione finora
- Fisiologia Del Liquido AmnioticoDocumento10 pagineFisiologia Del Liquido AmnioticoDanTe D' WinchesterNessuna valutazione finora
- Placenta Embriologia VeterinariaDocumento9 paginePlacenta Embriologia VeterinariaMoreno MoralesNessuna valutazione finora
- Hipoglicemia NeonatalDocumento12 pagineHipoglicemia NeonatalJuan Pablo AguilNessuna valutazione finora
- Atencion de Parto PDFDocumento21 pagineAtencion de Parto PDFrosmery supa floresNessuna valutazione finora
- Embarazo MultipleDocumento21 pagineEmbarazo MultipleJoanna VanessaNessuna valutazione finora
- 4-Fisiologia Del Trabajo de PartoDocumento20 pagine4-Fisiologia Del Trabajo de PartoOmar YesithNessuna valutazione finora
- Corazon EmbrionarioDocumento9 pagineCorazon EmbrionarioJhonatan Peña AcasieteNessuna valutazione finora
- Monografia de Gine OficialDocumento28 pagineMonografia de Gine OficialMarian GutierrezNessuna valutazione finora
- Enfermedad Trofoblastica GestacionalDocumento4 pagineEnfermedad Trofoblastica GestacionalYuritza AreliNessuna valutazione finora
- Barrera HematoencefálicaDocumento2 pagineBarrera HematoencefálicaGregory TejadaNessuna valutazione finora
- Embarazo en La AlturaDocumento8 pagineEmbarazo en La AlturaBrandonMendivilNessuna valutazione finora
- PDF - Clase de Inmunidad en El Feto y El NeonatoDocumento109 paginePDF - Clase de Inmunidad en El Feto y El Neonatokaren sanchezNessuna valutazione finora
- Polihidramniosy OligoamniosDocumento20 paginePolihidramniosy OligoamniosJolbelice ManceraNessuna valutazione finora
- Práctica Atención de PartoDocumento35 paginePráctica Atención de PartoRocio Dominguez MarureNessuna valutazione finora
- Vol20 3 1952 4Documento21 pagineVol20 3 1952 4Marco Julcamoro AsencioNessuna valutazione finora
- InmunidadDocumento14 pagineInmunidadJuanita Velasquez Zavala100% (1)
- Farmacoterapia A Lo Largo de La VidaDocumento15 pagineFarmacoterapia A Lo Largo de La Vidaana mariaNessuna valutazione finora
- Analgesia y Anestesia - InformeDocumento21 pagineAnalgesia y Anestesia - InformeMejia Vilchez LennanNessuna valutazione finora
- 14 Vitalidad Fetal Ecografia y DopplerDocumento5 pagine14 Vitalidad Fetal Ecografia y DopplerAgus PoncettaNessuna valutazione finora
- Embr 4 Placenta y Anexos Fetales MuniveDocumento38 pagineEmbr 4 Placenta y Anexos Fetales MuniveAndrés MesíaNessuna valutazione finora
- Endocrinología PlacentariaDocumento38 pagineEndocrinología PlacentariaLuis Aguagüiña ToaingaNessuna valutazione finora
- Sesión #5 - Cambios Anatomofisiológicos en El EmbarazoDocumento33 pagineSesión #5 - Cambios Anatomofisiológicos en El EmbarazoLuci Sánchez EspinozaNessuna valutazione finora
- Patología Periodontal y Parto Prematuro. Riesgo de Resultados Adversos Del EmbarazoDocumento15 paginePatología Periodontal y Parto Prematuro. Riesgo de Resultados Adversos Del EmbarazoJessica Izchel Velasco BarajasNessuna valutazione finora
- Destrezas Salvar VidasDocumento46 pagineDestrezas Salvar VidasVanessa RuthNessuna valutazione finora