Potrebbero piacerti anche
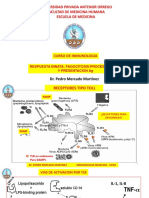
- Receptores Tipo TollDocumento5 pagineReceptores Tipo TollMayra Nedelka VegaNessuna valutazione finora
- Lectura de InmunidadDocumento8 pagineLectura de InmunidadKatherine Milagros Apcho VeraNessuna valutazione finora
- Mecanismos de Supresión de Las Células T Reguladoras (Treg) : ResumenDocumento9 pagineMecanismos de Supresión de Las Células T Reguladoras (Treg) : ResumenMariza100% (1)
- Fisiopatologia en Diabetes Mellitus 1Documento28 pagineFisiopatologia en Diabetes Mellitus 1Luis VerduzcoNessuna valutazione finora
- Capítulo I - Respuesta Inmune. de La Fisiología A La MedicinaDocumento26 pagineCapítulo I - Respuesta Inmune. de La Fisiología A La Medicinaicaro chaineNessuna valutazione finora
- DCM 183 LDocumento9 pagineDCM 183 LMARIN VALENCIA PAOLA ANDREANessuna valutazione finora
- Informe S11-Eayalas-101206 04M05-3Documento6 pagineInforme S11-Eayalas-101206 04M05-3Mafer GarcíaNessuna valutazione finora
- Fisiologia Del Sistema InmuneDocumento41 pagineFisiologia Del Sistema InmuneAndrea Lizeth Guerra JimenezNessuna valutazione finora
- TCRT (Transgenico)Documento14 pagineTCRT (Transgenico)Rosa BruckmannNessuna valutazione finora
- 4 Sistema Inmune Innato - IIDocumento18 pagine4 Sistema Inmune Innato - IIJ. JoseNessuna valutazione finora
- Universidad Privada Antenor Orrego Facultad de Medicina Humana Escuela de MedicinaDocumento19 pagineUniversidad Privada Antenor Orrego Facultad de Medicina Humana Escuela de MedicinaMonica García GutierrezNessuna valutazione finora
- 3 MicrobiotaDocumento6 pagine3 MicrobiotaMet LuNessuna valutazione finora
- CitokinasDocumento12 pagineCitokinasvanessa cañete garcíaNessuna valutazione finora
- Anestésicos en Sepsis. Conceptos ActualesDocumento8 pagineAnestésicos en Sepsis. Conceptos Actualesvelovi29103977Nessuna valutazione finora
- Bases Farmacológicas y Tratamiento de La InflamaciónDocumento50 pagineBases Farmacológicas y Tratamiento de La InflamaciónShabelNessuna valutazione finora
- Endotelio y SepsisDocumento9 pagineEndotelio y Sepsissilvia XNessuna valutazione finora
- EM y Las Celulas TDocumento9 pagineEM y Las Celulas Talbert gallardoNessuna valutazione finora
- Far Maco Genetic ADocumento11 pagineFar Maco Genetic AMimix NovedadNessuna valutazione finora
- Tecnicas de Infiltración IDocumento32 pagineTecnicas de Infiltración ICristiam Cuarezma100% (2)
- Quimioterapia y Trasplanet de Médula Ósea (12 y 13 Noviembre) - Presentación Del ProfesorDocumento80 pagineQuimioterapia y Trasplanet de Médula Ósea (12 y 13 Noviembre) - Presentación Del Profesorapi-3705495100% (3)
- Fisiopatologia de La SepsisDocumento21 pagineFisiopatologia de La Sepsisositto100% (1)
- LNKDocumento10 pagineLNKKary CocomNessuna valutazione finora
- 1er ParcialDocumento453 pagine1er ParcialMaria Fernanda Padron RiosNessuna valutazione finora
- Clase 0 IMDocumento33 pagineClase 0 IMDenisse PersonNessuna valutazione finora
- Actividad Receptores de PRRDocumento11 pagineActividad Receptores de PRRAndres PelayoNessuna valutazione finora
- Lectura Endotoxinas Su Importancia SalDocumento8 pagineLectura Endotoxinas Su Importancia SalOscarcito BrizuelaNessuna valutazione finora
- EmbejecimientoDocumento5 pagineEmbejecimientoJULIAN ENRRIQUE UTRIA BASTOSNessuna valutazione finora
- Tolerancia Periférica en Linfocito T, Tolerancia Periférica en Linfocito BDocumento6 pagineTolerancia Periférica en Linfocito T, Tolerancia Periférica en Linfocito BBryan Sancan50% (2)
- 7.enfermedades ReumaticasDocumento12 pagine7.enfermedades ReumaticasCarlos DecenaNessuna valutazione finora
- Inmunidad Innata Chuluyán ESP PDFDocumento20 pagineInmunidad Innata Chuluyán ESP PDFElías Damián Martínez TerénNessuna valutazione finora
- rr133d PDFDocumento8 paginerr133d PDFdaniloNessuna valutazione finora
- In Flamo SomaDocumento11 pagineIn Flamo SomaDuneska GomezNessuna valutazione finora
- Sepsis MaternaDocumento40 pagineSepsis MaternaAlberto BrunoNessuna valutazione finora
- Mecanismos Accion Farmacos InmunosupresoresDocumento16 pagineMecanismos Accion Farmacos InmunosupresoresDiana BenitesNessuna valutazione finora
- Regulación Molecular de La Polarización de La Respuesta InmuneDocumento9 pagineRegulación Molecular de La Polarización de La Respuesta InmuneVerónica Alejandra Pérez MoralesNessuna valutazione finora
- Epigenetica y Epigenomica de La Artritis ReumatoidDocumento14 pagineEpigenetica y Epigenomica de La Artritis ReumatoidXavi Martinez SantamariaNessuna valutazione finora
- P1 CirugíaDocumento59 pagineP1 CirugíaKelle MoralesNessuna valutazione finora
- Resumen InmunoDocumento71 pagineResumen InmunoGabriela OchoaNessuna valutazione finora
- Innate Immune Responses To Trauma - En.esDocumento15 pagineInnate Immune Responses To Trauma - En.esSalvador Jimenez MagosNessuna valutazione finora
- InflamacionDocumento3 pagineInflamacionDanielaNessuna valutazione finora
- 01 Introduccion A La HomotoxicologiaDocumento42 pagine01 Introduccion A La HomotoxicologiavicvansNessuna valutazione finora
- Cosas VariasDocumento9 pagineCosas VariasCarlos Espinosa MorenoNessuna valutazione finora
- Células de La Inmunidad Innata y Adaptativa en La Diabetes Mellitus Tipo 2 y ObesidadDocumento9 pagineCélulas de La Inmunidad Innata y Adaptativa en La Diabetes Mellitus Tipo 2 y ObesidadGiancarlo LindoNessuna valutazione finora
- Introduccion Al Sistema InmuneDocumento10 pagineIntroduccion Al Sistema InmuneAngelica AlvarezNessuna valutazione finora
- Respuesta Inmunitaria en El TraumaDocumento8 pagineRespuesta Inmunitaria en El TraumaYerlin Huamán BautistaNessuna valutazione finora
- Asp-Ap-R1-T7 (Parte 1)Documento25 pagineAsp-Ap-R1-T7 (Parte 1)Adrián Hyu ShisayNessuna valutazione finora
- Human Autoimmune Diseases A Comprehensive Update - En.esDocumento27 pagineHuman Autoimmune Diseases A Comprehensive Update - En.esNaye AponteNessuna valutazione finora
- Articulo Th1 y Th2Documento17 pagineArticulo Th1 y Th2Jesús Del ToroNessuna valutazione finora
- Semana 1-19 IiDocumento20 pagineSemana 1-19 IiElias Venancio BurgosNessuna valutazione finora
- Semana 2 - InmunologíaDocumento35 pagineSemana 2 - InmunologíaJosé Enrique Osorio De La CruzNessuna valutazione finora
- Artículo-La Respuesta Inmune AntiviralDocumento6 pagineArtículo-La Respuesta Inmune AntiviralGuillermo Alfonso Solórzano MoralesNessuna valutazione finora
- Mapa Conceptual La Artritis ReumatoideDocumento2 pagineMapa Conceptual La Artritis Reumatoidejpps0% (1)
- 11-Suplementación Alimentaria Con Algas Pardas - Efectos Sobre La Alergia y La Inflamación y Sus Consecuencias - PMCDocumento51 pagine11-Suplementación Alimentaria Con Algas Pardas - Efectos Sobre La Alergia y La Inflamación y Sus Consecuencias - PMC楊許志Nessuna valutazione finora
- ElsevierDocumento6 pagineElsevierpreguntando25Nessuna valutazione finora
- Dibnotas 1P CXDocumento125 pagineDibnotas 1P CXGerardo SanchezNessuna valutazione finora
- Alteraciones Del Sistema InmuneDocumento4 pagineAlteraciones Del Sistema InmuneAtex 11Nessuna valutazione finora
- Citoquinas en DermatologiaDocumento6 pagineCitoquinas en DermatologiaSile SciosciaNessuna valutazione finora
- Enf CeliacaDocumento17 pagineEnf CeliacaMomo HernándezNessuna valutazione finora
- Celulas LangerhansDocumento9 pagineCelulas Langerhansquelita123100% (5)
- RC004-ME-ET-AI-00-ID-U2-9002-00 Bombas para Cámaras de Coagulación - APCDocumento31 pagineRC004-ME-ET-AI-00-ID-U2-9002-00 Bombas para Cámaras de Coagulación - APCSergioNessuna valutazione finora
- Cuestionario Termografia Unidad IV 50 PreguntasDocumento11 pagineCuestionario Termografia Unidad IV 50 PreguntasALEJANDRO ENRIQUEZ FLORESNessuna valutazione finora
- TJJTNDocumento4 pagineTJJTNJosue GasparNessuna valutazione finora
- Efecto de Diferentes Configuraciones de Torque en La Formación de Grietas en La Dentina RadicularDocumento3 pagineEfecto de Diferentes Configuraciones de Torque en La Formación de Grietas en La Dentina Radicularyul garcia100% (1)
- Informe EspañolDocumento23 pagineInforme EspañolyoniNessuna valutazione finora
- Claudia Eliana SEGUNDO ENTREGABLEDocumento15 pagineClaudia Eliana SEGUNDO ENTREGABLEFranciscoAlmendrosGonzálezNessuna valutazione finora
- Papilomatosis Laringea RevisionDocumento10 paginePapilomatosis Laringea RevisionDavid Cáceres100% (1)
- UntitledDocumento2 pagineUntitledDarleny GutierrezNessuna valutazione finora
- Adicción Al Sexo PDFDocumento6 pagineAdicción Al Sexo PDFAnonymous SFtQd4zNessuna valutazione finora
- Reparacion de Nariz LeporinaDocumento12 pagineReparacion de Nariz LeporinaFabian Andres Portilla MogollonNessuna valutazione finora
- IntoxicaciónDocumento121 pagineIntoxicaciónCesar Britos AquinoNessuna valutazione finora
- Frutas y VerdurasDocumento10 pagineFrutas y Verdurasrosa guerreroNessuna valutazione finora
- ITB - 2021 - 21400 - FRM - CSI - ST - 0029 Check List ApisionadorDocumento1 paginaITB - 2021 - 21400 - FRM - CSI - ST - 0029 Check List ApisionadorrodolfoNessuna valutazione finora
- Programa Nacional de Saneamiento Rural (UE-005-001443)Documento64 paginePrograma Nacional de Saneamiento Rural (UE-005-001443)FRANCISCO GALLEGOS GARCIANessuna valutazione finora
- Capitulo IIDocumento40 pagineCapitulo IIariel sanchezNessuna valutazione finora
- I1 GenDocumento1 paginaI1 GenAlexandre Mendes100% (1)
- Costos de Producción en PorciculturaDocumento7 pagineCostos de Producción en PorciculturaEfrain DiazNessuna valutazione finora
- Elaboración de Néctar de Frutas TropicalesDocumento19 pagineElaboración de Néctar de Frutas TropicalesKatheryne Paola OchoaNessuna valutazione finora
- Patologías Del Sistema NerviosoDocumento68 paginePatologías Del Sistema NerviosoIvan GarciaNessuna valutazione finora
- Tema 11 - Patologías en EdificacionesDocumento23 pagineTema 11 - Patologías en EdificacionesLuis Lopez GonzalesNessuna valutazione finora
- Modelos SistémicosDocumento45 pagineModelos SistémicosVictor Alvarez MartinezNessuna valutazione finora
- Ejemplos de Aplicaciones TermosolarDocumento2 pagineEjemplos de Aplicaciones TermosolarRichard TatamuésNessuna valutazione finora
- La Relación de La Ética Con Las Ciencias Que Estudia El Comportamiento HumanoDocumento5 pagineLa Relación de La Ética Con Las Ciencias Que Estudia El Comportamiento HumanoAngel SaraviaNessuna valutazione finora
- FFDocumento12 pagineFFCAMILONessuna valutazione finora
- Mitos y Leyendas de Chimá Córdoba Geraldine Durante ColónDocumento3 pagineMitos y Leyendas de Chimá Córdoba Geraldine Durante ColónYANELSI HERNANDEZ CASTILLONessuna valutazione finora
- Guía de Estudio Trauma Segunda UnidadDocumento88 pagineGuía de Estudio Trauma Segunda UnidadKevinNessuna valutazione finora
- 1 Baterias Clases ElectricidadDocumento45 pagine1 Baterias Clases ElectricidadImirvlad GINessuna valutazione finora
- Seguro FacultativoDocumento2 pagineSeguro FacultativoEmporium CounterNessuna valutazione finora
- Resistencia Termica Por Contacto (Resumen)Documento6 pagineResistencia Termica Por Contacto (Resumen)Luis Alberto Aguayo Torrez100% (1)
- MatenimientoDocumento144 pagineMatenimientowilliamsNessuna valutazione finora