Potrebbero piacerti anche
- Enemigos de La MaderaDocumento44 pagineEnemigos de La MaderaEliiza AyalaNessuna valutazione finora
- Identificación Del Tipo de Pudrición de La Madera y Hongos Xilófagos en Árboles UrbanosDocumento9 pagineIdentificación Del Tipo de Pudrición de La Madera y Hongos Xilófagos en Árboles UrbanosEduardo Torres100% (1)
- 2.2 Degradacion de La MaderaDocumento41 pagine2.2 Degradacion de La MaderaEdwin Calderon100% (1)
- Gran Guía Visual para El Diagnóstico de Enfermedades en Las Plantas PDFDocumento36 pagineGran Guía Visual para El Diagnóstico de Enfermedades en Las Plantas PDFANDRES TAYLORNessuna valutazione finora
- Microorganismosdelsuelo 130513214147 Phpapp01Documento41 pagineMicroorganismosdelsuelo 130513214147 Phpapp01yeimirisNessuna valutazione finora
- Suelo Urbano y Los Vecinos InvisiblesDocumento18 pagineSuelo Urbano y Los Vecinos InvisiblesangelNessuna valutazione finora
- Ciclo de NitrógenoDocumento7 pagineCiclo de NitrógenoURIEL ALEJANDRO PADILLA MARTINEZNessuna valutazione finora
- Resumen BiomecanicaDocumento22 pagineResumen BiomecanicaMiluska SantosNessuna valutazione finora
- 3-Clase Arquitectura de Los Árboles 2020Documento35 pagine3-Clase Arquitectura de Los Árboles 2020AfterImageNessuna valutazione finora
- Tema 6 BotánicaDocumento88 pagineTema 6 Botánicaclara noelia martinez ramosNessuna valutazione finora
- I Parte Fito Cap I, II, IIIDocumento39 pagineI Parte Fito Cap I, II, IIIJMac ACNessuna valutazione finora
- INTERACCIONES DE LOS MICROORGANISMOS Y LA RAIZ DEL ARBOL URBANO (Autoguardado)Documento39 pagineINTERACCIONES DE LOS MICROORGANISMOS Y LA RAIZ DEL ARBOL URBANO (Autoguardado)Miguel SierraNessuna valutazione finora
- Sistema RadicularDocumento20 pagineSistema RadicularyesicolNessuna valutazione finora
- Raiz Tallo y HojasDocumento13 pagineRaiz Tallo y HojasCarmen GonzalezNessuna valutazione finora
- El TalloDocumento20 pagineEl TalloDany LuzaNessuna valutazione finora
- La Planta y Sus PartesDocumento9 pagineLa Planta y Sus PartesAriana PuiconNessuna valutazione finora
- Deteccion y Evaluacion de Defectos Peligrosos en Arboles PDFDocumento69 pagineDeteccion y Evaluacion de Defectos Peligrosos en Arboles PDFAlf AlvNessuna valutazione finora
- E-Hongos FitopatogenosDocumento22 pagineE-Hongos FitopatogenosAbel AlonsoNessuna valutazione finora
- Desarrollo SostenibleDocumento2 pagineDesarrollo SostenibleSelireth Reyes100% (1)
- BosqueDocumento15 pagineBosqueArelis EspinozaNessuna valutazione finora
- Morfología de HojaDocumento45 pagineMorfología de HojaIliana Pardo RojasNessuna valutazione finora
- Silvicultura I Unidad II. El Árbol 2023Documento32 pagineSilvicultura I Unidad II. El Árbol 2023Ruben PérezNessuna valutazione finora
- TEMA 7 - PaleobotánicaDocumento6 pagineTEMA 7 - PaleobotánicaAndrés ChapapoteNessuna valutazione finora
- Informe Identificacion Plagas de CitricosDocumento15 pagineInforme Identificacion Plagas de CitricosVERYK MOREYNessuna valutazione finora
- Actividad de Aprendizaje 4 PDFDocumento13 pagineActividad de Aprendizaje 4 PDFGabriel FernandezNessuna valutazione finora
- F Piojo BlancoDocumento2 pagineF Piojo BlancoJesúsNessuna valutazione finora
- Fitoplasmas y ProtozoariosDocumento8 pagineFitoplasmas y ProtozoariosPachoGarciaNessuna valutazione finora
- Ecología de InsectosDocumento40 pagineEcología de InsectosKarina Entoperú100% (1)
- Ciclo Del Carbono Materia Orgánica y HumusDocumento4 pagineCiclo Del Carbono Materia Orgánica y HumusJuan GuevaraNessuna valutazione finora
- SustratosDocumento8 pagineSustratosrosaburonNessuna valutazione finora
- Gestión Del Arbolado Urbano en EuropaDocumento66 pagineGestión Del Arbolado Urbano en EuropaJuan Manuel Sandin PérezNessuna valutazione finora
- Enfermedades de GeraniosDocumento2 pagineEnfermedades de Geraniospalomaazul2000Nessuna valutazione finora
- Fitopatologia General 2019 - IDocumento51 pagineFitopatologia General 2019 - IAnonymous ljIHRDNessuna valutazione finora
- PulgonesDocumento3 paginePulgonesInmaculadavanessa CasanovaNessuna valutazione finora
- Manual de Practicas FitopatologiaDocumento56 pagineManual de Practicas FitopatologiaRoberto Clemente Aguirre ElizondoNessuna valutazione finora
- Fertilizacion y Manejo Integrado de Plagas y EnfermedadesDocumento36 pagineFertilizacion y Manejo Integrado de Plagas y EnfermedadesJou AbreuNessuna valutazione finora
- FitonematologíaDocumento38 pagineFitonematologíacarlosmanuelacosta100% (1)
- Estructura y Desarrollo Del ArbolDocumento69 pagineEstructura y Desarrollo Del ArbolNINONessuna valutazione finora
- Manejo Agronomico Convencional Mango ManilaDocumento17 pagineManejo Agronomico Convencional Mango ManilaArianna BalamNessuna valutazione finora
- Enfermedades Vasculares Causadas Por HongosDocumento14 pagineEnfermedades Vasculares Causadas Por HongosTony TarazonaNessuna valutazione finora
- Factores Abioticos FisicosDocumento59 pagineFactores Abioticos FisicosGEANPIERRE SANGAY CACHINessuna valutazione finora
- 3-4 Uni EntomologiaDocumento313 pagine3-4 Uni EntomologiaFiorela VillalobosNessuna valutazione finora
- Plagas de La CebollaDocumento23 paginePlagas de La CebollaErnesto RosalesNessuna valutazione finora
- Pinus Caribaea PDFDocumento6 paginePinus Caribaea PDFMAGDA YULIETH DAZA MUNOZNessuna valutazione finora
- Manejo de Enfermedades BacteriasDocumento98 pagineManejo de Enfermedades BacteriasEvensJeanNessuna valutazione finora
- Hongo MildiuDocumento13 pagineHongo Mildiublackcat89Nessuna valutazione finora
- Fisiología de Plantas en Condiciones de EstrésDocumento21 pagineFisiología de Plantas en Condiciones de EstrésJeanpieer BG100% (1)
- MAGNOLIACEAEDocumento5 pagineMAGNOLIACEAETania SaloméNessuna valutazione finora
- Poda Arbolado Urbano y RosalesDocumento56 paginePoda Arbolado Urbano y RosalesmatíasNessuna valutazione finora
- Apuntes de FruticulturaDocumento18 pagineApuntes de FruticulturaOscar Pérez SaavedraNessuna valutazione finora
- Cultivo de MacroalgasDocumento12 pagineCultivo de Macroalgasyvet_951100% (7)
- Ciclos de Nutrientes de Las PlantasDocumento39 pagineCiclos de Nutrientes de Las PlantasHuarauyaGeronimoAdrianoNessuna valutazione finora
- Prueba de GerminaciónDocumento27 paginePrueba de GerminaciónFernando Valentin Bolaño PachecoNessuna valutazione finora
- Morfologia de CitricosDocumento9 pagineMorfologia de Citricosyeltsin rivera pinchiNessuna valutazione finora
- Otra 612 CalabacínDocumento20 pagineOtra 612 CalabacínSergi PillacaNessuna valutazione finora
- Árbol Colonial Frente A Árbol Unitario. Francis HalléDocumento0 pagineÁrbol Colonial Frente A Árbol Unitario. Francis HalléAtxe PreikestolenNessuna valutazione finora
- Tipos de Pudricion de La MaderaDocumento33 pagineTipos de Pudricion de La MaderaHector GarciaNessuna valutazione finora
- Hongos Xilófagos MONOGRAFIA PRESERVACIONDocumento6 pagineHongos Xilófagos MONOGRAFIA PRESERVACIONJuan Fernando Suescun BuitragoNessuna valutazione finora
- Pudrición de Los Árboles Fustales y Árboles Maduros en PieDocumento2 paginePudrición de Los Árboles Fustales y Árboles Maduros en Piedieguicho xeaNessuna valutazione finora
- Clases PrimariaDocumento4 pagineClases PrimariaMilagros Quiñones MarrerosNessuna valutazione finora
- El Cultivo Del MembrilloDocumento14 pagineEl Cultivo Del MembrilloJose Luis CANessuna valutazione finora
- ALGODONDocumento2 pagineALGODONMilagros Quiñones MarrerosNessuna valutazione finora
- Cartilla de AlcachofaDocumento2 pagineCartilla de AlcachofaMilagros Quiñones MarrerosNessuna valutazione finora
- CapsicumDocumento2 pagineCapsicumMilagros Quiñones MarrerosNessuna valutazione finora
- CapsicumDocumento2 pagineCapsicumMilagros Quiñones MarrerosNessuna valutazione finora
- Clase 3 Necesidades de Agua de Cultivos) Semmana 4Documento24 pagineClase 3 Necesidades de Agua de Cultivos) Semmana 4Milagros Quiñones MarrerosNessuna valutazione finora
- Teoria OfertaDocumento12 pagineTeoria OfertaMilagros Quiñones MarrerosNessuna valutazione finora
- Parte 4 - MacrosDocumento6 pagineParte 4 - MacrosMilagros Quiñones MarrerosNessuna valutazione finora
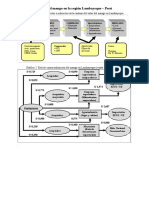
- La Cadena de Valor Del Mango en La Región LambayequeDocumento1 paginaLa Cadena de Valor Del Mango en La Región LambayequeMilagros Quiñones MarrerosNessuna valutazione finora
- Mercado AgrarioDocumento21 pagineMercado AgrarioMilagros Quiñones MarrerosNessuna valutazione finora
- Marketing y Plan de MarketinghgDocumento45 pagineMarketing y Plan de MarketinghgMilagros Quiñones MarrerosNessuna valutazione finora
- Teoria DemandaDocumento17 pagineTeoria DemandaMilagros Quiñones MarrerosNessuna valutazione finora
- Clase 4 Diseño de Riego Por Goteo (Sabado) Semana 4Documento38 pagineClase 4 Diseño de Riego Por Goteo (Sabado) Semana 4Milagros Quiñones MarrerosNessuna valutazione finora
- Encuesta de VermicompostDocumento10 pagineEncuesta de VermicompostMilagros Quiñones MarrerosNessuna valutazione finora
- Marchitez Bacteriana PDFDocumento25 pagineMarchitez Bacteriana PDFCadena Peruana MifarNessuna valutazione finora
- Marchitez Bacteriana PDFDocumento25 pagineMarchitez Bacteriana PDFCadena Peruana MifarNessuna valutazione finora
- 13 MIPgloboderapapaDocumento50 pagine13 MIPgloboderapapaMilagros Quiñones MarrerosNessuna valutazione finora
- Plagas de La Papa SierraDocumento87 paginePlagas de La Papa SierraYuri CalleNessuna valutazione finora
- Control Ecologico Articulo5Documento25 pagineControl Ecologico Articulo5040256Nessuna valutazione finora
- Cuestionario Sobre Los Gansos.Documento3 pagineCuestionario Sobre Los Gansos.Milagros Quiñones MarrerosNessuna valutazione finora
- Escuela Superior Politécnica de Chimborazo: Facultad de Informática Y ElectrónicaDocumento122 pagineEscuela Superior Politécnica de Chimborazo: Facultad de Informática Y ElectrónicaMilagros Quiñones MarrerosNessuna valutazione finora
- Marketing y Plan de MarketinghgDocumento45 pagineMarketing y Plan de MarketinghgMilagros Quiñones MarrerosNessuna valutazione finora
- 13 MIPgloboderapapaDocumento50 pagine13 MIPgloboderapapaMilagros Quiñones MarrerosNessuna valutazione finora
- Papa PDFDocumento68 paginePapa PDFOner Mendoza VasquezNessuna valutazione finora
- MangoDocumento2 pagineMangoMilagros Quiñones MarrerosNessuna valutazione finora
- Calculo de MaterialesDocumento3 pagineCalculo de MaterialesDavid RodriguezNessuna valutazione finora
- Algodon - VinsosDocumento2 pagineAlgodon - VinsosMilagros Quiñones MarrerosNessuna valutazione finora
- Cultivo de TOMATEDocumento4 pagineCultivo de TOMATEMilagros Quiñones MarrerosNessuna valutazione finora
- CapsicumDocumento2 pagineCapsicumMilagros Quiñones MarrerosNessuna valutazione finora
- CAPÍTULO 42 - Hongos. Conceptos BásicosDocumento12 pagineCAPÍTULO 42 - Hongos. Conceptos BásicosLESLIE LOPEZNessuna valutazione finora
- Parte 2. Biotecnología en La Agricultura. Cebio Ecuador.Documento145 pagineParte 2. Biotecnología en La Agricultura. Cebio Ecuador.Nelver MorenoNessuna valutazione finora
- Hongos ReproducciónDocumento43 pagineHongos ReproducciónAna Medaly MdccNessuna valutazione finora
- 1-Enfermedades de La MaderaDocumento13 pagine1-Enfermedades de La MaderaLORDMACLEOD15Nessuna valutazione finora
- Ejercicios Tema 6 La Clasificacic3b3n de Los Seres Vivos MicroorganismosDocumento3 pagineEjercicios Tema 6 La Clasificacic3b3n de Los Seres Vivos MicroorganismosCENSONessuna valutazione finora
- Inta Elaboracion de Pate PDFDocumento29 pagineInta Elaboracion de Pate PDFJessy MoriitzNessuna valutazione finora
- Taller de EcologiaDocumento2 pagineTaller de EcologiaPablo Salvador Calderon Santander100% (1)
- Laboratorio N°3 de SanidadDocumento9 pagineLaboratorio N°3 de SanidadAnonymous 3PE4OXfNessuna valutazione finora
- 20130816biofertilizantes Micorricicos FinalDocumento86 pagine20130816biofertilizantes Micorricicos FinalJuan CarlosNessuna valutazione finora
- Paracoccidioides BrasiliensisDocumento11 pagineParacoccidioides BrasiliensisvanessaNessuna valutazione finora
- Album Basidiomycota Grupo 4Documento118 pagineAlbum Basidiomycota Grupo 4DANNA DANEB NORIEGA REALESNessuna valutazione finora
- Patologia en La MaderaDocumento10 paginePatologia en La MaderaManuel Suare'zNessuna valutazione finora
- Guía 2 Estudio de Las Caracteristicas Macroscópicas y Microscópicas de Los Mohos y LevadurasDocumento6 pagineGuía 2 Estudio de Las Caracteristicas Macroscópicas y Microscópicas de Los Mohos y LevadurasLina Marcela Quintero VargasNessuna valutazione finora
- Abortos MicoticosDocumento18 pagineAbortos MicoticosVicente VegaNessuna valutazione finora
- Literatura Italiana Tp. Italo CalvinoDocumento8 pagineLiteratura Italiana Tp. Italo CalvinoarilaiseNessuna valutazione finora
- Nutricion Autotrofa PDFDocumento6 pagineNutricion Autotrofa PDFOrlando PayaresNessuna valutazione finora
- Hongos y LevadurasDocumento4 pagineHongos y LevadurasHannDy AlDairNessuna valutazione finora
- Manual Enfermedades&Pplagas AlfalfaDocumento64 pagineManual Enfermedades&Pplagas AlfalfaCesar Diaz Barrios50% (2)
- Informe Laboratorio de Biologia n1Documento16 pagineInforme Laboratorio de Biologia n1valen-escobarNessuna valutazione finora
- Cuadernillo de Actividades D. El 24 Al 27 de EneroDocumento20 pagineCuadernillo de Actividades D. El 24 Al 27 de EneroRomán Luna RosaNessuna valutazione finora
- ArchaebacteriaDocumento3 pagineArchaebacteriaalexNessuna valutazione finora
- Produccion de AlcoholDocumento10 pagineProduccion de AlcoholPiero PaoloNessuna valutazione finora
- Practica n2 Arce 11Documento9 paginePractica n2 Arce 11Luis Felipe Arce DiazNessuna valutazione finora
- Oídio en RosasDocumento15 pagineOídio en RosasParmenio Zuñiga IzquierdoNessuna valutazione finora
- INFORME 5. Torres MitziDocumento10 pagineINFORME 5. Torres MitziMITZI DAYANI TORRES MURILLONessuna valutazione finora
- Producción y Aplicación de TrichodermaDocumento7 pagineProducción y Aplicación de TrichodermaMara SatoNessuna valutazione finora
- Las Plantas 192569 - Libro-Al - 008-021 - Ud01 - Savia.Documento14 pagineLas Plantas 192569 - Libro-Al - 008-021 - Ud01 - Savia.Anderson Luis Cordero MateoNessuna valutazione finora
- Reino FungiDocumento13 pagineReino FungiGabriela Hernandez VargasNessuna valutazione finora
- Informe Manejo PoscosechaDocumento7 pagineInforme Manejo PoscosechaFabian Andres Rodriguez CastroNessuna valutazione finora
- RÍO MARÍA AGUILAR Inventario Flora y FaunaDocumento68 pagineRÍO MARÍA AGUILAR Inventario Flora y FaunaFernando Cabezas PraviaNessuna valutazione finora