Potrebbero piacerti anche
- DBS HuntingtonDocumento10 pagineDBS HuntingtonEdith SteinNessuna valutazione finora
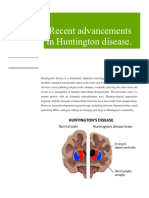
- Huntington's Disease: A Clinical ReviewDocumento32 pagineHuntington's Disease: A Clinical ReviewDewi NofiantiNessuna valutazione finora
- Huntington Disease Research PaperDocumento36 pagineHuntington Disease Research PaperJingyun WuNessuna valutazione finora
- PIIS0021925820492476Documento16 paginePIIS0021925820492476Roger MundiNessuna valutazione finora
- "Huntington Disease: Clinical Features and Diagnosis" "Huntington Disease: Management"Documento36 pagine"Huntington Disease: Clinical Features and Diagnosis" "Huntington Disease: Management"Gilbert LimenNessuna valutazione finora
- Huntington PDFDocumento11 pagineHuntington PDFMadalina Ionela StanciuNessuna valutazione finora
- Pilhatsch 2011Documento7 paginePilhatsch 2011omar rodriguezNessuna valutazione finora
- High and Low Levels of An NTRK2-Driven Genetic Profile Affect Motor - and Cognition-Associated Frontal Gray Matter in Prodromal Huntington's DiseaseDocumento24 pagineHigh and Low Levels of An NTRK2-Driven Genetic Profile Affect Motor - and Cognition-Associated Frontal Gray Matter in Prodromal Huntington's DiseaseJennifer CiarochiNessuna valutazione finora
- Reviews: Normal Huntingtin Function: An Alternative Approach To Huntington'S DiseaseDocumento12 pagineReviews: Normal Huntingtin Function: An Alternative Approach To Huntington'S DiseaseDaniel SchmidtNessuna valutazione finora
- Apuntes Caso ClínicoDocumento9 pagineApuntes Caso ClínicoViviana LunaNessuna valutazione finora
- 7,8-Dihydroxyflavone, A Small Molecular TRKB Agonist, Is Useful For Treating Various Bdnf-Implicated Human DisordersDocumento9 pagine7,8-Dihydroxyflavone, A Small Molecular TRKB Agonist, Is Useful For Treating Various Bdnf-Implicated Human DisordersJ.J.Nessuna valutazione finora
- Huntington’s Disease: Pathogenic Mechanisms and Implications for TherapeuticsDa EverandHuntington’s Disease: Pathogenic Mechanisms and Implications for TherapeuticsX. William YangNessuna valutazione finora
- Use of MTHFR C677T Polymorphism and Plasma Pharmacokinetics To Predict Methotrexate Toxicity in Patients With Acute Lymphoblastic LeukemiaDocumento8 pagineUse of MTHFR C677T Polymorphism and Plasma Pharmacokinetics To Predict Methotrexate Toxicity in Patients With Acute Lymphoblastic LeukemiaFaMe TanapornNessuna valutazione finora
- 2018 - Rol de La MITOCONDRIA y HUNTINGTONDocumento25 pagine2018 - Rol de La MITOCONDRIA y HUNTINGTONEduardo RodriguezNessuna valutazione finora
- 1 s2.0 S1773224721007024 MainDocumento34 pagine1 s2.0 S1773224721007024 MainNihal ChauhanNessuna valutazione finora
- Huntingtons Treatment PDFDocumento8 pagineHuntingtons Treatment PDFave123izationNessuna valutazione finora
- Moléculas PaperDocumento44 pagineMoléculas PaperCarlla Araújo Guarani KaiowáNessuna valutazione finora
- Glutamate-Modulating Drugs As A Potential Therapeutic Strategy in Obsessive-Compulsive DisorderDocumento18 pagineGlutamate-Modulating Drugs As A Potential Therapeutic Strategy in Obsessive-Compulsive DisorderjuliermetostaNessuna valutazione finora
- Paper 8 SidiquiDocumento31 paginePaper 8 SidiquiNICOLE HSIAO-SANCHEZNessuna valutazione finora
- Bayram-Weston Et Al 2016 - MHTT Antibodies in Mouse ModelsDocumento15 pagineBayram-Weston Et Al 2016 - MHTT Antibodies in Mouse ModelsJingyun WuNessuna valutazione finora
- Anti Inflammatory Effects of New Human Histamine H3 Receptor Receptor Ligands With Flavonoid Structure On BV-2 NeuroinflammationDocumento14 pagineAnti Inflammatory Effects of New Human Histamine H3 Receptor Receptor Ligands With Flavonoid Structure On BV-2 NeuroinflammationEwelina HonkiszNessuna valutazione finora
- EZH2: Not EZHY (Easy) To Deal: ReviewDocumento16 pagineEZH2: Not EZHY (Easy) To Deal: ReviewLorena RamosNessuna valutazione finora
- HadDocumento20 pagineHadRiddhi GandhiNessuna valutazione finora
- Resveratrol Attenuates Methamphetamine-Induced Memory Impairment Via Inhibition of Oxidative Stress and Apoptosis in MiceDocumento15 pagineResveratrol Attenuates Methamphetamine-Induced Memory Impairment Via Inhibition of Oxidative Stress and Apoptosis in MiceMohammad Hadi SahebiNessuna valutazione finora
- Menzies 2015Documento13 pagineMenzies 2015kosikevinonuNessuna valutazione finora
- OT6 - Huntington's DiseaseDocumento1 paginaOT6 - Huntington's DiseaseAnnbe BarteNessuna valutazione finora
- Glioblastoma Growth and Invasiveness Receptor Channel TRPC6 Is A Key Mediator of Notch-DrivenDocumento13 pagineGlioblastoma Growth and Invasiveness Receptor Channel TRPC6 Is A Key Mediator of Notch-DrivenPlia NattineeNessuna valutazione finora
- Review HNDocumento44 pagineReview HNLeilane GlienkeNessuna valutazione finora
- Azadiradione Restores Protein Quality Control and Ameliorates The Disease Pathogenesis in A Mouse Model of Huntington 'S DiseaseDocumento10 pagineAzadiradione Restores Protein Quality Control and Ameliorates The Disease Pathogenesis in A Mouse Model of Huntington 'S DiseaseAnirban MannaNessuna valutazione finora
- Modeling Alzheimer's...Documento35 pagineModeling Alzheimer's...clfp25Nessuna valutazione finora
- Advances in The PharmacologicalDocumento12 pagineAdvances in The PharmacologicalpaijosusenoNessuna valutazione finora
- Mutant Huntingtin Impairs Axonal Trafficking in MaDocumento16 pagineMutant Huntingtin Impairs Axonal Trafficking in MaAnders AskersrudNessuna valutazione finora
- PsychFest - Lipid Droplets in HepatocytesDocumento1 paginaPsychFest - Lipid Droplets in HepatocytesAj KeefeNessuna valutazione finora
- 2001 Gray and Roth Brain Research BulletinDocumento11 pagine2001 Gray and Roth Brain Research BulletinBrener Santos Da SilvaNessuna valutazione finora
- BV2 Publikacja Final Version 06.02.22 For SendingDocumento11 pagineBV2 Publikacja Final Version 06.02.22 For SendingEwelina HonkiszNessuna valutazione finora
- The Phenomenal Epigenome in NeurodevelopmentalDocumento9 pagineThe Phenomenal Epigenome in Neurodevelopmentaleup_1983Nessuna valutazione finora
- Huntingtons DiseaseDocumento3 pagineHuntingtons DiseaseKavya tripathiNessuna valutazione finora
- Huntingtons ChoreaDocumento11 pagineHuntingtons ChoreajananiNessuna valutazione finora
- Giannaccini2013 The Expression of Platelet Serotonin Transporter (SERT) in Human Obesity.Documento8 pagineGiannaccini2013 The Expression of Platelet Serotonin Transporter (SERT) in Human Obesity.ArizonaNessuna valutazione finora
- s41380 021 01384 8Documento10 pagines41380 021 01384 8mariamojegNessuna valutazione finora
- HDAC3 Inhibition Up Regulates PD-L1 Expression in B-Cell Lymphomas and Augments The Efficacy of Anti-PD-L1 TherapyDocumento28 pagineHDAC3 Inhibition Up Regulates PD-L1 Expression in B-Cell Lymphomas and Augments The Efficacy of Anti-PD-L1 Therapyshidis1028Nessuna valutazione finora
- The Tale of Histone Modifications and Its Role in Multiple SclerosisDocumento12 pagineThe Tale of Histone Modifications and Its Role in Multiple SclerosisMercedes del Pilar Canchihuamán SuasnábarNessuna valutazione finora
- Journal Pcbi 1010708Documento28 pagineJournal Pcbi 1010708hilaire0902Nessuna valutazione finora
- Emerging Epigenetic-Modulating - Sermer2019Documento14 pagineEmerging Epigenetic-Modulating - Sermer2019Diego AntunezNessuna valutazione finora
- Chen 2018Documento11 pagineChen 2018Mohamed El-AgamyNessuna valutazione finora
- Dissertation HdacDocumento4 pagineDissertation HdacWillYouWriteMyPaperForMeAlbuquerque100% (1)
- Choline Transporter Gene Variation Is Associated With Attention-Deficit Hyperactivity DisorderDocumento12 pagineCholine Transporter Gene Variation Is Associated With Attention-Deficit Hyperactivity DisordervikNessuna valutazione finora
- Hun A Clini 2022 AndhaleDocumento7 pagineHun A Clini 2022 AndhaleAngelica Maria PardoNessuna valutazione finora
- Gut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersDocumento10 pagineGut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersVitor PereiraNessuna valutazione finora
- Gut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersDocumento10 pagineGut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersLori EsbeltoNessuna valutazione finora
- Dasko, 2022Documento33 pagineDasko, 2022farihameetsworldNessuna valutazione finora
- MYT1L Haploinsufficiency in Human NeuronsDocumento14 pagineMYT1L Haploinsufficiency in Human Neuronsredmi karimNessuna valutazione finora
- HDACDocumento10 pagineHDACfarihameetsworldNessuna valutazione finora
- Cancers 13 04088 v3Documento18 pagineCancers 13 04088 v3xjwwm99v6nNessuna valutazione finora
- InmunohistoquímicaDocumento18 pagineInmunohistoquímicaViviana LunaNessuna valutazione finora
- Ihv 2018 Poster-FinalDocumento1 paginaIhv 2018 Poster-Finalapi-447486080Nessuna valutazione finora
- 494-Vortioxetine 5HTDocumento7 pagine494-Vortioxetine 5HTamiralbertoNessuna valutazione finora
- 2015 - Hyperthermina, An Effective Strategy To Induce Apoptosis in Cancer CellsDocumento9 pagine2015 - Hyperthermina, An Effective Strategy To Induce Apoptosis in Cancer CellsAndrea MolinaNessuna valutazione finora
- Quinderé Et Al 2015 PDFDocumento9 pagineQuinderé Et Al 2015 PDFLuizaNessuna valutazione finora
- Histo NervousDocumento20 pagineHisto NervousALESSANDRA CELINessuna valutazione finora
- A Cultured Affair HSV Latency and Reactivation in Neurons Jan2014Documento8 pagineA Cultured Affair HSV Latency and Reactivation in Neurons Jan2014Ana-Maria DavidNessuna valutazione finora
- Alternative MedicineDocumento226 pagineAlternative Medicinegvrnaidu3200Nessuna valutazione finora
- Connections Between Mathematics and Biology: Carl C. Cowen I Dept of Mathematical SciencesDocumento42 pagineConnections Between Mathematics and Biology: Carl C. Cowen I Dept of Mathematical Sciencesjais_sankarNessuna valutazione finora
- Neurotransmitters NotesDocumento5 pagineNeurotransmitters NotesLIZNessuna valutazione finora
- Anatomy of The Sympathetic Nervous SystemDocumento12 pagineAnatomy of The Sympathetic Nervous SystemNanda 9300Nessuna valutazione finora
- Zygmunt L. Kruk, Christopher J. Pycock (Auth.) - Neurotransmitters and Drugs-Springer Netherlands (1991)Documento210 pagineZygmunt L. Kruk, Christopher J. Pycock (Auth.) - Neurotransmitters and Drugs-Springer Netherlands (1991)joaomartinelliNessuna valutazione finora
- Mindmap Bio621 Chapter1Documento3 pagineMindmap Bio621 Chapter1MizahNessuna valutazione finora
- Axonal Transport (Neuron 2014)Documento35 pagineAxonal Transport (Neuron 2014)Eduardo RamirezNessuna valutazione finora
- Vanders Chapter 6Documento53 pagineVanders Chapter 6gablimlikescheeseNessuna valutazione finora
- NBME 19 BlockDocumento201 pagineNBME 19 BlockHaider Ali100% (3)
- Mitocondria y ParkinsonDocumento13 pagineMitocondria y ParkinsonEstebanNessuna valutazione finora
- Peripheral Nerve Problems: Anne M.B. MosconyDocumento40 paginePeripheral Nerve Problems: Anne M.B. MosconyAkbar GazaliNessuna valutazione finora
- Nerve PhysiologyDocumento23 pagineNerve PhysiologyGladdyll Raico DizonNessuna valutazione finora
- M1-M6 Biochem LabDocumento383 pagineM1-M6 Biochem Labloreign sinocruzNessuna valutazione finora
- Lecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without NotesDocumento119 pagineLecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without Notesellie marcusNessuna valutazione finora
- Mental and Elemental NutrientsDocumento21 pagineMental and Elemental NutrientsGilbert RivaNessuna valutazione finora
- Test Bank For Foundations of Behavioral Neuroscience Paper 9 e 9th Edition Neil R CarlsonDocumento26 pagineTest Bank For Foundations of Behavioral Neuroscience Paper 9 e 9th Edition Neil R CarlsonPaula Smith100% (5)
- Introduction To Neuroscience 1702495272Documento789 pagineIntroduction To Neuroscience 1702495272LauNessuna valutazione finora
- Fundamentals of Nervous System and Nervous TissueDocumento37 pagineFundamentals of Nervous System and Nervous Tissue차지형Nessuna valutazione finora
- Histology of Nervous TissueDocumento40 pagineHistology of Nervous TissueSHARON MARIA SUNNYNessuna valutazione finora
- Physiology U-3 Excitable Nervous TissueDocumento122 paginePhysiology U-3 Excitable Nervous Tissuesinte beyuNessuna valutazione finora
- On-Line Assignment DefinitionsDocumento2 pagineOn-Line Assignment DefinitionsAbbey AdlerNessuna valutazione finora
- Cytoskeleton PDFDocumento48 pagineCytoskeleton PDFDave RapaconNessuna valutazione finora
- Anatomy of The Human Optic Nerve: Structure and Function: Provisional ChapterDocumento20 pagineAnatomy of The Human Optic Nerve: Structure and Function: Provisional ChapterAndres BenavidesNessuna valutazione finora
- Ganong All MCQDocumento55 pagineGanong All MCQSagor Kumar DasNessuna valutazione finora
- Peripheral Neuropathies in Clinical PracticeDocumento398 paginePeripheral Neuropathies in Clinical PracticeIka Gultom100% (1)