Potrebbero piacerti anche
- Digestion y Absorcion de NutrientesDocumento15 pagineDigestion y Absorcion de NutrientesAspirante Folio 530313393Nessuna valutazione finora
- Absorcio IntestinalDocumento4 pagineAbsorcio IntestinalPiero LlanosNessuna valutazione finora
- Fisiología DigestivaDocumento84 pagineFisiología DigestivaRICARDO ALBERTO GUTIÉRREZ ABARZÚANessuna valutazione finora
- Taller de Nutricion FinalDocumento11 pagineTaller de Nutricion FinalJhoan David Londoño MendozaNessuna valutazione finora
- Intestino Grueso Monogástricos y RumiantesDocumento5 pagineIntestino Grueso Monogástricos y RumiantesSeba OrellanaNessuna valutazione finora
- Clase 22 Sistema Digestivo PDFDocumento26 pagineClase 22 Sistema Digestivo PDFQuimica TecnologiaNessuna valutazione finora
- Proteinas, Proteinas Animales y VegetalesDocumento9 pagineProteinas, Proteinas Animales y VegetalesRamé TMNessuna valutazione finora
- Conceptualizacion y Clasificacion de Los AlimentosDocumento5 pagineConceptualizacion y Clasificacion de Los Alimentosyulitza milena miranda cuadro100% (1)
- Tarea Digestión: LogíaDocumento4 pagineTarea Digestión: LogíaEmily Reyes100% (1)
- Boca Estómago Intestino Delgado Nutrientes Resultantes: 5.-Completa La Siguiente TablaDocumento2 pagineBoca Estómago Intestino Delgado Nutrientes Resultantes: 5.-Completa La Siguiente TablaEmily ReyesNessuna valutazione finora
- 3 Nutricion y DietaDocumento18 pagine3 Nutricion y DietaHENRY JESUS GONZALES SOSANessuna valutazione finora
- Digestión y Absorción de NutrientesDocumento27 pagineDigestión y Absorción de NutrientesGinna SolarteNessuna valutazione finora
- Principios De Nutrición De RumiantesDa EverandPrincipios De Nutrición De RumiantesNessuna valutazione finora
- BLOQUE 1 - MÓDULO 1 - CLASE 1 - Nutrición Clínica I Bases Moleculares de La NutriciónDocumento7 pagineBLOQUE 1 - MÓDULO 1 - CLASE 1 - Nutrición Clínica I Bases Moleculares de La NutriciónArantxa BernasconiNessuna valutazione finora
- Teoria Macro y MicronutrientesDocumento14 pagineTeoria Macro y MicronutrientesGuadeloupe PalNessuna valutazione finora
- CLASE 36-Fisiología de La Digestión Del Intestino Delgado, Digestión Mecánica y Química.Documento46 pagineCLASE 36-Fisiología de La Digestión Del Intestino Delgado, Digestión Mecánica y Química.SHERLY DEL CARMEN HUANCAS OCUPANessuna valutazione finora
- Flora Intestinal PDFDocumento5 pagineFlora Intestinal PDFEder Soncco BravoNessuna valutazione finora
- Fisiología Gastrointestinal TripticoDocumento2 pagineFisiología Gastrointestinal TripticoGrace Garcia100% (1)
- Taller Digestion Nutricion B22Documento5 pagineTaller Digestion Nutricion B228bmorenoaguirrejuanestebanNessuna valutazione finora
- Resumen Nutricion AnimalDocumento7 pagineResumen Nutricion AnimalMaría del Mar CNessuna valutazione finora
- Alimentos y Nutrientes Parte 1Documento48 pagineAlimentos y Nutrientes Parte 1Paloma LópezNessuna valutazione finora
- DigestivDocumento19 pagineDigestivLeo SimbañaNessuna valutazione finora
- Taller BioquimicaDocumento6 pagineTaller BioquimicaValentina NuñezNessuna valutazione finora
- Sist. Digestivo y Metabolismo OKDocumento7 pagineSist. Digestivo y Metabolismo OKDianna ValentinNessuna valutazione finora
- Taller - Segundo Corte - Nutricion - A2020Documento13 pagineTaller - Segundo Corte - Nutricion - A2020DANIEL SANCHEZ100% (1)
- Los Péptidos Bioactivos en AlimentaciónDocumento5 pagineLos Péptidos Bioactivos en AlimentaciónAraceli NolascoNessuna valutazione finora
- Importancia de Los Carbohidratos en La Alimentación BovinaDocumento6 pagineImportancia de Los Carbohidratos en La Alimentación BovinaCarlos Orlando Palma Oliva100% (1)
- Microbiota y ProbioticosDocumento56 pagineMicrobiota y ProbioticosgquilNessuna valutazione finora
- Infografía - Importancia de La Nutrición para La Salud y El Rendimiento FísicoDocumento1 paginaInfografía - Importancia de La Nutrición para La Salud y El Rendimiento FísicoEDGARDO SÁNCHEZ MARTINEZNessuna valutazione finora
- Informe de Tracto Digestivo..Documento6 pagineInforme de Tracto Digestivo..Javier Alejandro Vizcarra GonzalesNessuna valutazione finora
- 4B Act 2-Cytabs. Nut.Documento2 pagine4B Act 2-Cytabs. Nut.Kevan Etyan Rivera QuevedoNessuna valutazione finora
- Desarrollo Nutricional de La ComunidadDocumento6 pagineDesarrollo Nutricional de La ComunidadAlejandra Emilin Rojas RamirezNessuna valutazione finora
- Rol de La Fibra DietariaDocumento9 pagineRol de La Fibra DietariaJuliana TovarNessuna valutazione finora
- El Metabolismo Lipídico Bovino y Su Relación Con La DietaDocumento11 pagineEl Metabolismo Lipídico Bovino y Su Relación Con La DietaGary Jheferson Salazar RondonNessuna valutazione finora
- Proteina ArticuloDocumento5 pagineProteina ArticuloSusan ChinoNessuna valutazione finora
- Exposición Jardon, Et Al.Documento14 pagineExposición Jardon, Et Al.Juan Gonzalez FernandezNessuna valutazione finora
- El Aparato Digestivo para Tercero de SecundariaDocumento8 pagineEl Aparato Digestivo para Tercero de SecundariaJoshua Monroe100% (3)
- Análisis de Artículo 2 - Ácido EstomacalDocumento3 pagineAnálisis de Artículo 2 - Ácido EstomacalEstefania FrancoNessuna valutazione finora
- Semana 6 - Teoria BioquimicaDocumento56 pagineSemana 6 - Teoria BioquimicaLuciana AronesNessuna valutazione finora
- Microbiota Intestinal ScribdDocumento2 pagineMicrobiota Intestinal ScribdAndrea CalderonNessuna valutazione finora
- InterrelacionesDocumento24 pagineInterrelacionesEmery Sofía C. D. BustamanteNessuna valutazione finora
- Tema II Cheng 2020Documento14 pagineTema II Cheng 2020Luis Ángel Rod FloNessuna valutazione finora
- NUTRICION Y ALIMENTACION EN CUY Presentacion 1Documento20 pagineNUTRICION Y ALIMENTACION EN CUY Presentacion 1Alex Pan de GraciaNessuna valutazione finora
- Fisiopatología y Mecanismos de Acción de Los Prebióticos - MedwaveDocumento6 pagineFisiopatología y Mecanismos de Acción de Los Prebióticos - MedwaveXochitl PonceNessuna valutazione finora
- Digestión, Absorción y Asimilación de CarbohidratosDocumento33 pagineDigestión, Absorción y Asimilación de CarbohidratosNoreidys Julieth Arias Romero100% (2)
- Unidad 7 Introduccion Al Metabolismo I - 521Documento4 pagineUnidad 7 Introduccion Al Metabolismo I - 521Carolina Longoria SNessuna valutazione finora
- Anatomía Gastrointestinal Del ConejoDocumento27 pagineAnatomía Gastrointestinal Del ConejoMiguel SotoNessuna valutazione finora
- Profundizacion ESIC ESFUNO Digestivo PDFDocumento27 pagineProfundizacion ESIC ESFUNO Digestivo PDFEvelyn FENFNessuna valutazione finora
- Tema 7Documento9 pagineTema 7Suárez CíoNessuna valutazione finora
- Alimentos Prebióticos y ProbióticosDocumento6 pagineAlimentos Prebióticos y ProbióticosMartha_ArNessuna valutazione finora
- Teoría - Semana 11Documento20 pagineTeoría - Semana 11matt enmeNessuna valutazione finora
- Fibra Dietética - Resumen de Artículo - Mota ArmandoDocumento3 pagineFibra Dietética - Resumen de Artículo - Mota ArmandoArmando OlivaresNessuna valutazione finora
- Mapa ConceptualDocumento1 paginaMapa ConceptualJhon Perez PerezNessuna valutazione finora
- Clase 7. Farmacocinética ExcreciónDocumento40 pagineClase 7. Farmacocinética ExcreciónVanessa ortiz GuevaraNessuna valutazione finora
- Función Del Intestino GruesoDocumento5 pagineFunción Del Intestino GruesoIsa Cruz HipolitoNessuna valutazione finora
- Taller Digestion Nutricion b2021Documento8 pagineTaller Digestion Nutricion b2021Katherine Perdomo RondonNessuna valutazione finora
- Digestion de GrasasDocumento9 pagineDigestion de GrasasYaritsa Tapia VasquezNessuna valutazione finora
- Informe Determinación de Fibra PDFDocumento4 pagineInforme Determinación de Fibra PDFEnrique MoraNessuna valutazione finora
- Sistema Digestivo 2020 PDFDocumento34 pagineSistema Digestivo 2020 PDFLina Cordero BerrioNessuna valutazione finora
- 046ger PDFDocumento56 pagine046ger PDFalicia dawnNessuna valutazione finora
- Proyecto Final Metodo EquipoCherryDocumento23 pagineProyecto Final Metodo EquipoCherryalicia dawnNessuna valutazione finora
- Comunicación Nutriólogo - PacienteDocumento23 pagineComunicación Nutriólogo - Pacientealicia dawnNessuna valutazione finora
- Módulo 1 Tarea 2Documento2 pagineMódulo 1 Tarea 2alicia dawnNessuna valutazione finora
- Cirrosis Del Libro CecilDocumento5 pagineCirrosis Del Libro Cecilalicia dawnNessuna valutazione finora
- AakDocumento2 pagineAakalicia dawnNessuna valutazione finora
- Capítulo IiiDocumento84 pagineCapítulo Iiialicia dawnNessuna valutazione finora
- Biometría HemáticaDocumento14 pagineBiometría Hemáticaalicia dawn0% (1)
- Lo Que Conmunmente Se Llama Antropologia Cognitiva EtnosemanticaDocumento10 pagineLo Que Conmunmente Se Llama Antropologia Cognitiva Etnosemanticaalicia dawnNessuna valutazione finora
- CIRUGIA - TegumentoDocumento38 pagineCIRUGIA - TegumentoVicente Mendoza JiménezNessuna valutazione finora
- Escala de Signos Blandos326Documento6 pagineEscala de Signos Blandos326Laura Roa AlonsoNessuna valutazione finora
- Clasificación TaxonomicaDocumento16 pagineClasificación Taxonomicayoseli moralesNessuna valutazione finora
- Infecciones Bacterianas Primarias de Piel y Tejidos BlandosDocumento8 pagineInfecciones Bacterianas Primarias de Piel y Tejidos BlandossteffanyNessuna valutazione finora
- Baloncesto Cuestionario EspolDocumento6 pagineBaloncesto Cuestionario EspolAdolfo VillaquiránNessuna valutazione finora
- Presiones Inspiratoria y Espiratoria MaximaDocumento28 paginePresiones Inspiratoria y Espiratoria MaximaYessenia Aranda OlguinNessuna valutazione finora
- FolletoDocumento16 pagineFolletoZully CabreraNessuna valutazione finora
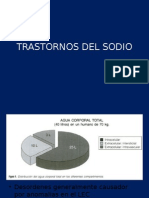
- Trastornos Del Sodio FisiopatologíaDocumento31 pagineTrastornos Del Sodio FisiopatologíaErick HernandezNessuna valutazione finora
- Consolidación Trabajo de Toxicologia Fase 5Documento18 pagineConsolidación Trabajo de Toxicologia Fase 5Luz Karina Pestana CaballeroNessuna valutazione finora
- Tu Felicidad Depende de Un PerroDocumento3 pagineTu Felicidad Depende de Un PerroChris CarvajalNessuna valutazione finora
- Protocolos Resonanca Magnética MaiteDocumento22 pagineProtocolos Resonanca Magnética MaiteMarco Mendez GonzalezNessuna valutazione finora
- Cuídate Compa! - Manual de La Autogestión de La Salud - Salud AntiautoritariaDocumento80 pagineCuídate Compa! - Manual de La Autogestión de La Salud - Salud AntiautoritariamilenasnNessuna valutazione finora
- Tanatologia EscritoDocumento19 pagineTanatologia Escrito1804147559Nessuna valutazione finora
- Plantilla TareaDocumento5 paginePlantilla Tareahector0% (1)
- Ficha Excreción AnimalesDocumento1 paginaFicha Excreción AnimalesCiencias Naturales SSNessuna valutazione finora
- Practica 17. Tejido y Sistema LinfoideDocumento9 paginePractica 17. Tejido y Sistema LinfoideJavier Osorio100% (1)
- Sist-Nervioso Estructura y Funcion Esquema AlumnosDocumento1 paginaSist-Nervioso Estructura y Funcion Esquema AlumnosRocío Mora Mora GomezNessuna valutazione finora
- Ejemplo de Ficha TextualDocumento3 pagineEjemplo de Ficha Textualjorgechipanavega0% (1)
- Oestrus OvisDocumento1 paginaOestrus Ovisyelko donairesNessuna valutazione finora
- Unamirada 590Documento1 paginaUnamirada 590Leon Peña RamosNessuna valutazione finora
- BIOQUÍMICA DENTAL Harvey - Bioquimica - 5e - Addendum PDFDocumento20 pagineBIOQUÍMICA DENTAL Harvey - Bioquimica - 5e - Addendum PDFbrayan emmanuel100% (1)
- 1.-EL NIÑO PREMATURO (Caracteristicas Anatomofuncionales)Documento38 pagine1.-EL NIÑO PREMATURO (Caracteristicas Anatomofuncionales)Jose David Gracia Martinez100% (2)
- Al Mal Paso Darle PrisaDocumento130 pagineAl Mal Paso Darle PrisaDANIELA CARDONA RAMIREZNessuna valutazione finora
- Enfermedades de La Piel EquinaDocumento2 pagineEnfermedades de La Piel EquinaAntonioTozkyNessuna valutazione finora
- I Resumen CamelloDocumento2 pagineI Resumen CamelloJose Martin Chuctaya MacedoNessuna valutazione finora
- Sistema EndocrinoDocumento39 pagineSistema EndocrinoYo Te CuidareNessuna valutazione finora
- Examen Médico Erick Estrada PDFDocumento2 pagineExamen Médico Erick Estrada PDFJulio Cesar Landeo FernandezNessuna valutazione finora
- Fecundacion Desarrollo Embrionario y FetalDocumento49 pagineFecundacion Desarrollo Embrionario y FetaljaigpameNessuna valutazione finora
- Síndrome Bi 2013Documento63 pagineSíndrome Bi 2013polilla74Nessuna valutazione finora
- Embarazo EctopicoDocumento14 pagineEmbarazo EctopicoAbraham MolinaNessuna valutazione finora