Potrebbero piacerti anche
- Taller de Limpieza Energetica Integral. OkDocumento40 pagineTaller de Limpieza Energetica Integral. OkIMRM6093% (15)
- Presentacion Sobre La Fermentacion AlcoholicaDocumento21 paginePresentacion Sobre La Fermentacion AlcoholicaKathy MeGo0% (1)
- Tecnología para la producción de cacao. Como sistema agroforestal en regiones potenciales de ColombiaDa EverandTecnología para la producción de cacao. Como sistema agroforestal en regiones potenciales de ColombiaNessuna valutazione finora
- Cinética de Procesos FermentativosDocumento7 pagineCinética de Procesos FermentativosJorge Lin YangNessuna valutazione finora
- Avances de la Biotecnología y su impacto en la agricultura latinoamericanaDa EverandAvances de la Biotecnología y su impacto en la agricultura latinoamericanaNessuna valutazione finora
- Cromatografia en PapelDocumento14 pagineCromatografia en PapelCamila Marmolejo VergaraNessuna valutazione finora
- Monografia de Levaduras.Documento12 pagineMonografia de Levaduras.Wilfredo Villanueva PuenteNessuna valutazione finora
- SEPU-MC-04 Calculo de Barra de 10 KVDocumento3 pagineSEPU-MC-04 Calculo de Barra de 10 KVErika Buenalaya CangalayaNessuna valutazione finora
- Vida Util ProductoDocumento23 pagineVida Util Productojulio h.oNessuna valutazione finora
- Guía Práctica 8 Fermentación AlcoholicaDocumento11 pagineGuía Práctica 8 Fermentación AlcoholicaJuli Osorio50% (4)
- Proceso de Fermentacion PDFDocumento8 pagineProceso de Fermentacion PDFPaco OlveraNessuna valutazione finora
- Proceso de AlineaciÓnDocumento6 pagineProceso de AlineaciÓnCésar Malo86% (21)
- Poi 2023 Atm-FpoDocumento33 paginePoi 2023 Atm-Fpojulio h.o100% (1)
- Alex - Roco - Tarea 2.Documento11 pagineAlex - Roco - Tarea 2.Alex Antonio Roco NeyraNessuna valutazione finora
- Cultivos Iniciadores Del VinoDocumento13 pagineCultivos Iniciadores Del VinoaraicaNessuna valutazione finora
- Acetobacter TrabajoDocumento10 pagineAcetobacter TrabajoGabby Naranjo Freire80% (5)
- ESCLAVISMODocumento11 pagineESCLAVISMOJesus BeltranNessuna valutazione finora
- Biotecnologia EnologicaDocumento4 pagineBiotecnologia EnologicaCristobal Hernandez ToledoNessuna valutazione finora
- Aislamiento de Levadura Productora de Etanol A Partir de Chicha de JoraDocumento4 pagineAislamiento de Levadura Productora de Etanol A Partir de Chicha de JoraKeylita GavidiaNessuna valutazione finora
- Fermentación Alcoholica Con LevadurasDocumento4 pagineFermentación Alcoholica Con LevadurasLaura Franco SalazarNessuna valutazione finora
- FermentaciónDocumento8 pagineFermentaciónJeyson GonzalezNessuna valutazione finora
- Biorefinerias TM - Clase X HE-fermentacion-biotecnologiaDocumento24 pagineBiorefinerias TM - Clase X HE-fermentacion-biotecnologiaHernan Felipe Cifuentes PeñaNessuna valutazione finora
- Aislamiento de Levaduras Productoras de Etanol (Pedro)Documento4 pagineAislamiento de Levaduras Productoras de Etanol (Pedro)Osmar Jarir Gonzales ZavaletaNessuna valutazione finora
- Fermentacion AlcoholicaDocumento3 pagineFermentacion AlcoholicaCarlos Cruz0% (2)
- Informe de Fermentación AlcoholicaDocumento19 pagineInforme de Fermentación AlcoholicaWily Jhonatan Bermeo Pérez0% (1)
- Trabajo de Biotecnologia (Vinos)Documento14 pagineTrabajo de Biotecnologia (Vinos)Melissa Nataly Huacasi QuispeNessuna valutazione finora
- PRÁCTICA No. 6Documento7 paginePRÁCTICA No. 6Estefania Rojas RayoNessuna valutazione finora
- Tipos de FermentacionDocumento7 pagineTipos de FermentacionLuis carlos Quiguantar chamorroNessuna valutazione finora
- Fermentacion de La LevaduraDocumento9 pagineFermentacion de La LevaduraCeciMoralesNessuna valutazione finora
- Aplicaciones de La Ingeniería Genética y La Genómica en EnologíaDocumento9 pagineAplicaciones de La Ingeniería Genética y La Genómica en EnologíaanaiNessuna valutazione finora
- Práctica 5Documento6 paginePráctica 5Valentina Buendia QuinteroNessuna valutazione finora
- Fermentación. IntroducciónDocumento26 pagineFermentación. Introducciónhenry molinaNessuna valutazione finora
- Fermentación en BatchDocumento3 pagineFermentación en BatchGerman DiazNessuna valutazione finora
- Efectos de La TemperaturaDocumento11 pagineEfectos de La TemperaturaDiego Quiroz AlborNessuna valutazione finora
- Zymomonas Móbilis ExpoDocumento4 pagineZymomonas Móbilis Exponacuc_22Nessuna valutazione finora
- Fermentacion Alcoholica de La Pomarrosa-Grupo 5Documento27 pagineFermentacion Alcoholica de La Pomarrosa-Grupo 5Brandon RodriguezNessuna valutazione finora
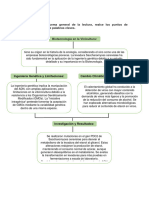
- Realice Un Esquema General de La Lectura, Realce Los Puntos de Importancia y Use Palabras ClavesDocumento2 pagineRealice Un Esquema General de La Lectura, Realce Los Puntos de Importancia y Use Palabras ClavesHeydi Graciela Albites AquijeNessuna valutazione finora
- Fermentacion Alcoholica VinosDocumento5 pagineFermentacion Alcoholica VinosRonald0% (1)
- Métodos Moleculares de Identificación de Microorganismos (1) - PDFDocumento10 pagineMétodos Moleculares de Identificación de Microorganismos (1) - PDFMartha BenavidesNessuna valutazione finora
- Aplicación de Un Bioproceso Industrial en El EcuadorDocumento8 pagineAplicación de Un Bioproceso Industrial en El EcuadorISAAC JORDY LLIVISUPA BRAVONessuna valutazione finora
- Principales Microorganismos Que Intervienen en La Fermentación AlcohólicaDocumento4 paginePrincipales Microorganismos Que Intervienen en La Fermentación AlcohólicaJoselynParedesNessuna valutazione finora
- Tercer ExamenDocumento10 pagineTercer ExamenVicente MendozaNessuna valutazione finora
- Fermentacion Alcoholica Del VinoDocumento7 pagineFermentacion Alcoholica Del VinoPaola Padilla BelmonteNessuna valutazione finora
- Evaluacion Del Proceso FermentativoDocumento3 pagineEvaluacion Del Proceso FermentativocarlosNessuna valutazione finora
- Proyecto BioprocesosDocumento12 pagineProyecto BioprocesosEdgar MaldonadoNessuna valutazione finora
- BioprocesosDocumento2 pagineBioprocesosGiovanny Jimenez MezaNessuna valutazione finora
- Bacterias Vino 2 PDFDocumento6 pagineBacterias Vino 2 PDFAnalia Díaz SaavedraNessuna valutazione finora
- Tecnica de Fermentacion AlcoholicaDocumento4 pagineTecnica de Fermentacion AlcoholicaPORTILLA PAGOLA RUBY DEL PILARNessuna valutazione finora
- Fermentacion AlcoholicaDocumento7 pagineFermentacion AlcoholicasusypaoNessuna valutazione finora
- PP Ten ZimasDocumento9 paginePP Ten ZimasAmir NeibNessuna valutazione finora
- Análisis Cinético de La Curva Básica de Crecimiento de Levaduras y de La Producción de EtanolDocumento5 pagineAnálisis Cinético de La Curva Básica de Crecimiento de Levaduras y de La Producción de EtanolMacias Guadalupe LizbethNessuna valutazione finora
- Informe FermentacionDocumento23 pagineInforme FermentacionMCamilaVillanuevaAvila100% (1)
- COFERMENTACIONESDocumento2 pagineCOFERMENTACIONESiredelvelNessuna valutazione finora
- El Uso de Saccharomyces Cerevisiae Como Bio-Fábrica para La Producción de BioetanolDocumento4 pagineEl Uso de Saccharomyces Cerevisiae Como Bio-Fábrica para La Producción de Bioetanolshaermmm124Nessuna valutazione finora
- Resuemen Etanol de Sorgo DulceDocumento6 pagineResuemen Etanol de Sorgo Dulcedaniela llanosNessuna valutazione finora
- Conservacion Por FermentacionDocumento40 pagineConservacion Por FermentacionHansNessuna valutazione finora
- Metodo de Fermentación Con Levadura Sccharomyces Cerevisiae para La Obtención de Bioetanol de Cáscara de NaranjaDocumento25 pagineMetodo de Fermentación Con Levadura Sccharomyces Cerevisiae para La Obtención de Bioetanol de Cáscara de Naranjaandres.pechoa0% (1)
- Cuestionario Laboratorio 6Documento2 pagineCuestionario Laboratorio 6Noelia Alondra MartinezNessuna valutazione finora
- 13 - Fase 2 Caracterización Del Proyecto.Documento11 pagine13 - Fase 2 Caracterización Del Proyecto.JACKELINE TORRES PEDREROSNessuna valutazione finora
- Fundamentos de La Fermentación Alcohólica: 1.-IntroduccionDocumento11 pagineFundamentos de La Fermentación Alcohólica: 1.-IntroduccionPaqui Alonso MingoranceNessuna valutazione finora
- Fermentación AlcohólicaDocumento27 pagineFermentación AlcohólicaJocelyn CristernaNessuna valutazione finora
- Informe Vino Fermentado Grupo 1Documento25 pagineInforme Vino Fermentado Grupo 1Irene VelascoNessuna valutazione finora
- MONOGRAFIA ActualDocumento18 pagineMONOGRAFIA ActualeliaNessuna valutazione finora
- Alimentos Geneticamente ModificadosDocumento12 pagineAlimentos Geneticamente ModificadosAlexandra Limache AlaveNessuna valutazione finora
- Conferencia 2 Conceptos Básicos Sobre FermentaciónDocumento13 pagineConferencia 2 Conceptos Básicos Sobre FermentaciónLore DelisNessuna valutazione finora
- Aislamiento de Microorganismo Tolerantes A EtanolDocumento6 pagineAislamiento de Microorganismo Tolerantes A EtanolCarolina GomezNessuna valutazione finora
- Tema 7. Microorganismos Fermentadores (Apuntes)Documento18 pagineTema 7. Microorganismos Fermentadores (Apuntes)XaviNessuna valutazione finora
- Acta de EntregaDocumento9 pagineActa de Entregajulio h.oNessuna valutazione finora
- Modelo Resolucion de Atm 2023Documento5 pagineModelo Resolucion de Atm 2023julio h.oNessuna valutazione finora
- Informe Atm 2023 - Felix PachecoDocumento85 pagineInforme Atm 2023 - Felix Pachecojulio h.oNessuna valutazione finora
- Ejercicio de Plan AgregadoDocumento1 paginaEjercicio de Plan Agregadojulio h.oNessuna valutazione finora
- DEBILIDADES InternasDocumento1 paginaDEBILIDADES Internasjulio h.oNessuna valutazione finora
- Plan de Trabajo Anual Muni Andarapa 2023Documento13 paginePlan de Trabajo Anual Muni Andarapa 2023julio h.oNessuna valutazione finora
- Ejemplos de Gestion Por ProcesoDocumento1 paginaEjemplos de Gestion Por Procesojulio h.oNessuna valutazione finora
- Municipalidad Centro Poblado PuyhuallaDocumento2 pagineMunicipalidad Centro Poblado Puyhuallajulio h.oNessuna valutazione finora
- Grupo - I - de Analisis e Interpretacion de DatosDocumento12 pagineGrupo - I - de Analisis e Interpretacion de Datosjulio h.oNessuna valutazione finora
- Comunidad ChantaDocumento3 pagineComunidad Chantajulio h.oNessuna valutazione finora
- Informe de Pia 2020 IIDocumento5 pagineInforme de Pia 2020 IIjulio h.oNessuna valutazione finora
- HistoriaDocumento2 pagineHistoriajulio h.oNessuna valutazione finora
- Las Ovejas TraduccionDocumento9 pagineLas Ovejas Traduccionjulio h.oNessuna valutazione finora
- PuyhuallaDocumento2 paginePuyhuallajulio h.oNessuna valutazione finora
- EXAMEN CapacidadesDocumento5 pagineEXAMEN Capacidadesjulio h.oNessuna valutazione finora
- DEBILIDADES InternasDocumento1 paginaDEBILIDADES Internasjulio h.oNessuna valutazione finora
- FotosDocumento1 paginaFotosjulio h.oNessuna valutazione finora
- Quilca PorrasDocumento290 pagineQuilca Porrasjulio h.oNessuna valutazione finora
- Control Grupos de 4Documento1 paginaControl Grupos de 4julio h.oNessuna valutazione finora
- Anexo #01Documento1 paginaAnexo #01julio h.oNessuna valutazione finora
- Tremistre de PuyhuallaDocumento1 paginaTremistre de Puyhuallajulio h.oNessuna valutazione finora
- Municipalidad Centro Poblado PuyhuallaDocumento2 pagineMunicipalidad Centro Poblado Puyhuallajulio h.oNessuna valutazione finora
- Metodologia de Teñido de CueroDocumento12 pagineMetodologia de Teñido de Cuerojulio h.oNessuna valutazione finora
- Diseño de Producto EPIADocumento19 pagineDiseño de Producto EPIAjulio h.oNessuna valutazione finora
- Fuente de Tension ReguladaDocumento2 pagineFuente de Tension Reguladajulio h.oNessuna valutazione finora
- Fuente de Tension ReguladaDocumento2 pagineFuente de Tension Reguladajulio h.oNessuna valutazione finora
- Diseño de Producto EPIADocumento19 pagineDiseño de Producto EPIAjulio h.oNessuna valutazione finora
- Solicitud Uso de VacacionesDocumento1 paginaSolicitud Uso de Vacacionesjulio h.oNessuna valutazione finora
- PEC4 - Marta BattistellaDocumento3 paginePEC4 - Marta BattistellaKevin SarmientoNessuna valutazione finora
- 480-Texto Del Artículo-1426-1-10-20160119Documento10 pagine480-Texto Del Artículo-1426-1-10-20160119Wenddy BarillasNessuna valutazione finora
- Caracas para PrincipiantesDocumento147 pagineCaracas para PrincipiantesSolángel RoccocuchiNessuna valutazione finora
- Milagro EucaristicoDocumento1 paginaMilagro EucaristicoRenzo GomezNessuna valutazione finora
- Bolet N Miski Mayo Marzo 2018Documento3 pagineBolet N Miski Mayo Marzo 2018Erick Daniel Rivera GómezNessuna valutazione finora
- Proyecto de Investigacion - Tecnologia NFC - EXPODocumento22 pagineProyecto de Investigacion - Tecnologia NFC - EXPOjetzelNessuna valutazione finora
- Vigilancia Epidemiológica y Vigilancia en La Salud PúblicaDocumento8 pagineVigilancia Epidemiológica y Vigilancia en La Salud Pública用户未知100% (1)
- INFORME PASANTIAS JAVIER VASQUEZ C.I 21.470.361 SECCION 4 Rev.Documento15 pagineINFORME PASANTIAS JAVIER VASQUEZ C.I 21.470.361 SECCION 4 Rev.javier vasquezNessuna valutazione finora
- Funciones y Modelos MatemáticosDocumento25 pagineFunciones y Modelos MatemáticosMario LNessuna valutazione finora
- Friedrichpaul Los Principes de NaranjaDocumento7 pagineFriedrichpaul Los Principes de NaranjabisitosNessuna valutazione finora
- Patron Datos Afovic 2019Documento38 paginePatron Datos Afovic 2019Aldo Lázaro RodriguezNessuna valutazione finora
- Leccion 1Documento2 pagineLeccion 1leonel21Nessuna valutazione finora
- Departamento de CaldasDocumento5 pagineDepartamento de CaldasLafachaga ChagaNessuna valutazione finora
- Atención y MemoriaDocumento31 pagineAtención y MemoriaJesus Smith Atocha CalleNessuna valutazione finora
- Actividades de La Unidad IV EspañolDocumento5 pagineActividades de La Unidad IV Españolsolterito1188Nessuna valutazione finora
- Clera 1683Documento15 pagineClera 1683Dra PeñaNessuna valutazione finora
- Actividad Individual Punto 3 Viviana Andrea GalindoDocumento4 pagineActividad Individual Punto 3 Viviana Andrea GalindovivianaNessuna valutazione finora
- Anexo 2 Tarea 3Documento2 pagineAnexo 2 Tarea 3kata morenoNessuna valutazione finora
- Maíz DuroDocumento3 pagineMaíz DuroJose PopNessuna valutazione finora
- Actividad Virtual N°4 - HidrologiaDocumento6 pagineActividad Virtual N°4 - HidrologiaJOSE ANGEL RONY ARAGON VALENCIANessuna valutazione finora
- Diseño de PozosDocumento23 pagineDiseño de PozosmiguelNessuna valutazione finora
- Plan2° 2012-2013Documento17 paginePlan2° 2012-2013Miguel Eduardo Sanchez Frias100% (1)
- Campos Consolidados y de Desarrollo Profesional de Los Ingenieros en MéxicoDocumento10 pagineCampos Consolidados y de Desarrollo Profesional de Los Ingenieros en MéxicoDaniela PerezNessuna valutazione finora
- Analitico Modificacion FebDocumento59 pagineAnalitico Modificacion FebLUIS FERNANDO APAZA YUCRANessuna valutazione finora