Potrebbero piacerti anche
- Down Syndrome and Incidental Pediatric Basal Ganglia Calcifications at AutopsyDocumento3 pagineDown Syndrome and Incidental Pediatric Basal Ganglia Calcifications at AutopsyPaul HartingNessuna valutazione finora
- Repulsive Lubrication On Hydrophilic SurfacesDocumento4 pagineRepulsive Lubrication On Hydrophilic SurfacesPaul HartingNessuna valutazione finora
- Structural and Functional Brain Differences in Key Opinion Journal LeadersDocumento4 pagineStructural and Functional Brain Differences in Key Opinion Journal LeadersPaul HartingNessuna valutazione finora
- Multifocal Tuberculosis in Southern Tunisia What Is Specific With ?Documento4 pagineMultifocal Tuberculosis in Southern Tunisia What Is Specific With ?Paul HartingNessuna valutazione finora
- Changes in The Structure of Synovial Fluids Between Healthy Joints and in OsteoarthritisDocumento3 pagineChanges in The Structure of Synovial Fluids Between Healthy Joints and in OsteoarthritisPaul HartingNessuna valutazione finora
- Crop Tolerance Against Salt StressDocumento2 pagineCrop Tolerance Against Salt StressPaul HartingNessuna valutazione finora
- Indications and Clinical Outcomes of High Tibial Osteotomy A Literature ReviewDocumento8 pagineIndications and Clinical Outcomes of High Tibial Osteotomy A Literature ReviewPaul HartingNessuna valutazione finora
- Aneurysmal Bone Cyst of The Fibular Malleolus Report of An Additional Case at Rare SiteDocumento4 pagineAneurysmal Bone Cyst of The Fibular Malleolus Report of An Additional Case at Rare SitePaul HartingNessuna valutazione finora
- Simultaneous Determination of The Ratio of Live, Dead and Adherent Bacteria in The Presence of Various Single-Walled Carbon NanotubesDocumento5 pagineSimultaneous Determination of The Ratio of Live, Dead and Adherent Bacteria in The Presence of Various Single-Walled Carbon NanotubesPaul HartingNessuna valutazione finora
- CAR-T Cell Immunotherapy and Nanotechnology: An Integrated Approach For Cancer CareDocumento2 pagineCAR-T Cell Immunotherapy and Nanotechnology: An Integrated Approach For Cancer CarePaul HartingNessuna valutazione finora
- Effect of Size On Gold Nanoparticles in Radiation Therapy Uptake and Survival EffectsDocumento8 pagineEffect of Size On Gold Nanoparticles in Radiation Therapy Uptake and Survival EffectsPaul HartingNessuna valutazione finora
- Molecular Analysis For Genetic Diversity and Population Structure of Ethiopian Faba Bean (Vicia Faba L) AccessionsDocumento10 pagineMolecular Analysis For Genetic Diversity and Population Structure of Ethiopian Faba Bean (Vicia Faba L) AccessionsPaul HartingNessuna valutazione finora
- Generation and Characterization of A Monoclonal Antibody Against Pseudorabies Virus Glycoprotein CDocumento5 pagineGeneration and Characterization of A Monoclonal Antibody Against Pseudorabies Virus Glycoprotein CPaul HartingNessuna valutazione finora
- Two Stage Treatment of Concomitant Complicated Right Amyand's Hernia and Incarcerated Left Inguinal Hernia Case Report and Review of The LiteratureDocumento3 pagineTwo Stage Treatment of Concomitant Complicated Right Amyand's Hernia and Incarcerated Left Inguinal Hernia Case Report and Review of The LiteraturePaul HartingNessuna valutazione finora
- Maternal and Fetal Effects of Cardiac Illness During Pregnancy in Beni Suef LocalitiesEgypt A Multi Centric Cross Sectional StudyDocumento7 pagineMaternal and Fetal Effects of Cardiac Illness During Pregnancy in Beni Suef LocalitiesEgypt A Multi Centric Cross Sectional StudyPaul HartingNessuna valutazione finora
- Examining The Association of Right Ventricular Dysfunction With Moderate To Severe Bronchopulmonary Dysplasia in Preterm InfantsDocumento7 pagineExamining The Association of Right Ventricular Dysfunction With Moderate To Severe Bronchopulmonary Dysplasia in Preterm InfantsPaul HartingNessuna valutazione finora
- Seborrheic Keratosis of The Breast Mimicking MelanomaDocumento3 pagineSeborrheic Keratosis of The Breast Mimicking MelanomaPaul HartingNessuna valutazione finora
- Pigmented Bowen Disease On Sclerosus and Atrophicus LichenDocumento3 paginePigmented Bowen Disease On Sclerosus and Atrophicus LichenPaul HartingNessuna valutazione finora
- Difference Between Being in Favor of Permitting Abortion After Rape and Willingness To Provide Such Services According To Physician's CharacteristicsDocumento4 pagineDifference Between Being in Favor of Permitting Abortion After Rape and Willingness To Provide Such Services According To Physician's CharacteristicsPaul HartingNessuna valutazione finora
- Extra Genital Lichen Sclero AtrophicDocumento2 pagineExtra Genital Lichen Sclero AtrophicPaul HartingNessuna valutazione finora
- Oral Apthous Ulcer Healing StepsDocumento2 pagineOral Apthous Ulcer Healing StepsPaul HartingNessuna valutazione finora
- Prevalence of Anti Cyclic Citrullinated Peptide Antibodies in Juvenile Idiopathic Arthritis in BeninDocumento4 paginePrevalence of Anti Cyclic Citrullinated Peptide Antibodies in Juvenile Idiopathic Arthritis in BeninPaul HartingNessuna valutazione finora
- Neuromyelitis Optica Spectrum Disorder With Spinal Involvement Including Conus Medullaris Lesions With Anti-Aquaporin-4 Antibody: A Pediatric Case and Literature ReviewDocumento3 pagineNeuromyelitis Optica Spectrum Disorder With Spinal Involvement Including Conus Medullaris Lesions With Anti-Aquaporin-4 Antibody: A Pediatric Case and Literature ReviewPaul HartingNessuna valutazione finora
- Who Was HakimDocumento2 pagineWho Was HakimPaul HartingNessuna valutazione finora
- A Novel Use of Extracorporeal Membrane Oxygenation in The Management of Prosthetic Valve Thrombosis During PregnancyDocumento4 pagineA Novel Use of Extracorporeal Membrane Oxygenation in The Management of Prosthetic Valve Thrombosis During PregnancyPaul HartingNessuna valutazione finora
- Recombinant Human Bone Morphogenetic Protein 2 For Correction of Unilateral Cleft Lip and Palatal Case ReportDocumento4 pagineRecombinant Human Bone Morphogenetic Protein 2 For Correction of Unilateral Cleft Lip and Palatal Case ReportPaul HartingNessuna valutazione finora
- Use of Short Term Orthodontics by The Specialist OrthodontistDocumento5 pagineUse of Short Term Orthodontics by The Specialist OrthodontistPaul HartingNessuna valutazione finora
- Factors Associated With Self Rated Oral Health in Adolescents in São Luís Maranhão Brazil 2014Documento8 pagineFactors Associated With Self Rated Oral Health in Adolescents in São Luís Maranhão Brazil 2014Paul HartingNessuna valutazione finora
- Benign Stromal Tumor Breast ImagesDocumento2 pagineBenign Stromal Tumor Breast ImagesPaul HartingNessuna valutazione finora
- Potassium Channels in Breast CancerDocumento6 paginePotassium Channels in Breast CancerPaul HartingNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
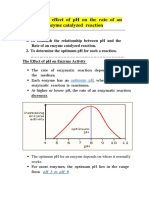
- Effect of PH On Enzyme ActivityDocumento12 pagineEffect of PH On Enzyme ActivityAb AbNessuna valutazione finora
- Anatomy and Physiology Chapter11 The Muscular SystemDocumento18 pagineAnatomy and Physiology Chapter11 The Muscular SystemDayledaniel SorvetoNessuna valutazione finora
- Fs 1 Episode 4 Learner Diversity PDF Learning RelationshipsDocumento1 paginaFs 1 Episode 4 Learner Diversity PDF Learning RelationshipsEmmarie Sola - FriersonNessuna valutazione finora
- Paired Passages - PresentationDocumento17 paginePaired Passages - Presentationsuchi ravaliaNessuna valutazione finora
- 2457-Article Text-14907-2-10-20120724Documento6 pagine2457-Article Text-14907-2-10-20120724desi meleniaNessuna valutazione finora
- Designing For The Human SensesDocumento50 pagineDesigning For The Human SensesRahaf MachaalNessuna valutazione finora
- Midterm 3TDocumento10 pagineMidterm 3TThanh JessieNessuna valutazione finora
- The Lab Session Report of Biological Techniques in Laboratory and Field (BIO403) Animals and Plants Colllection TechniquesDocumento5 pagineThe Lab Session Report of Biological Techniques in Laboratory and Field (BIO403) Animals and Plants Colllection TechniquesUmmi SalamahNessuna valutazione finora
- Lesson 3.1 History of Life On EarthDocumento1 paginaLesson 3.1 History of Life On EarthJoshua EboraNessuna valutazione finora
- Activity 2. Plant Cell Vs Animal CellDocumento5 pagineActivity 2. Plant Cell Vs Animal CellMary Joyce BonayonNessuna valutazione finora
- PPPGGGPPRRDocumento18 paginePPPGGGPPRRRully SetiawanNessuna valutazione finora
- Curs Ginkgo BilobaDocumento7 pagineCurs Ginkgo BilobaAnamaria RizacNessuna valutazione finora
- Ecosystems Thinking For Mind EcosystemsDocumento2 pagineEcosystems Thinking For Mind EcosystemsBala PillaiNessuna valutazione finora
- Competency Audit Grade 8Documento5 pagineCompetency Audit Grade 8Laurel S. MedinaNessuna valutazione finora
- Ujmr 2020 - 4 - 24 - 30 PDFDocumento7 pagineUjmr 2020 - 4 - 24 - 30 PDFUMYU Journal of Microbiology Research (UJMR)Nessuna valutazione finora
- ABO Blood Type and Personality Traits in Healthy Japanese SubjectsDocumento10 pagineABO Blood Type and Personality Traits in Healthy Japanese SubjectsDuta Nur HidayadNessuna valutazione finora
- FeatureSelectionAccepted IEEE Review PDFDocumento20 pagineFeatureSelectionAccepted IEEE Review PDFrvsamy80Nessuna valutazione finora
- Energy Pyramid - Activity StephanieDocumento4 pagineEnergy Pyramid - Activity Stephanieapi-473892313Nessuna valutazione finora
- Components of Physical FitnessDocumento1 paginaComponents of Physical Fitnessarjay zxcNessuna valutazione finora
- Biology Lab ReportDocumento13 pagineBiology Lab Reportapi-390838152Nessuna valutazione finora
- Northern Blot Analysis For Detection of RNA in PanDocumento8 pagineNorthern Blot Analysis For Detection of RNA in PanFati ZoraNessuna valutazione finora
- BRO Fluorescence SpectrosDocumento24 pagineBRO Fluorescence SpectrosWaleed El-azabNessuna valutazione finora
- Anima and Animus Term ReportDocumento17 pagineAnima and Animus Term ReportSadia Shahid100% (1)
- Genevieve Balunis Resume 2017Documento3 pagineGenevieve Balunis Resume 2017Dakota BalunisNessuna valutazione finora
- Effect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly PopulationDocumento6 pagineEffect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly Populationastrinila fauziNessuna valutazione finora
- General Biology - Chapter IDocumento10 pagineGeneral Biology - Chapter IG.k. Vinnan Rao100% (2)
- 8th Class Biology Project WorkDocumento10 pagine8th Class Biology Project WorkPRANU83% (6)
- The Effect of Temperature On The Hatching Success of Brine ShrimpDocumento2 pagineThe Effect of Temperature On The Hatching Success of Brine ShrimptahamidNessuna valutazione finora
- DKA Protocol TGDocumento11 pagineDKA Protocol TGabelNessuna valutazione finora
- Lewis Terman - Genius and StupidityDocumento78 pagineLewis Terman - Genius and StupiditymarnekibNessuna valutazione finora