Potrebbero piacerti anche
- Potencial de Reposo y EquilibrioDocumento5 paginePotencial de Reposo y EquilibrioDan PariascaNessuna valutazione finora
- Cilios y FlagelosDocumento6 pagineCilios y FlagelosXavi Zambrano GINessuna valutazione finora
- Sinapsis. Quimica y Electrica ppt4Documento8 pagineSinapsis. Quimica y Electrica ppt4Camilo San Martin CarrascoNessuna valutazione finora
- Sistema Muscular Del Cuerpo Humano y Sus PartesDocumento5 pagineSistema Muscular Del Cuerpo Humano y Sus PartesLaura CastilloNessuna valutazione finora
- URETRADocumento10 pagineURETRAcamachitaNessuna valutazione finora
- Fisiología NeuromuscularDocumento5 pagineFisiología NeuromuscularOscar Placencia AgredaNessuna valutazione finora
- Modelo Electrico de La Menbrana CelularDocumento8 pagineModelo Electrico de La Menbrana CelularAlex GarciaNessuna valutazione finora
- Pares Craneales y Su FuncionDocumento25 paginePares Craneales y Su FuncionRobinsonGiraldoLópez100% (1)
- Capitulo Señalizacion Celular-ReceptoresDocumento60 pagineCapitulo Señalizacion Celular-ReceptoresEdwar Leyva Baltazar0% (1)
- Intro Historia de Los AnestésicosDocumento12 pagineIntro Historia de Los AnestésicosGonzalo Paredes GonzalesNessuna valutazione finora
- Cuestionario de Tejido NerviosoDocumento2 pagineCuestionario de Tejido NerviosoAna Karen Aguilar RicoNessuna valutazione finora
- 03 - Alteraciones de La ConcienciaDocumento34 pagine03 - Alteraciones de La ConcienciaAle ZambranoNessuna valutazione finora
- Region CigomaticaDocumento35 pagineRegion CigomaticaAJ_evaNessuna valutazione finora
- Generacion y Conduccion de Potenciales de AcciónDocumento8 pagineGeneracion y Conduccion de Potenciales de AcciónIván Andrés Escobar Figueroa0% (1)
- Sistematización Anatómica de La Médula EspinalDocumento3 pagineSistematización Anatómica de La Médula EspinalVictorFloresAnchanteNessuna valutazione finora
- Histologia Del CorazonDocumento16 pagineHistologia Del Corazonlaura katalina mejia rodriguezNessuna valutazione finora
- Cap 47 Sensibilidades Somáticas - I. Organización General, Las Sensaciones Táctil y Posicional PDFDocumento8 pagineCap 47 Sensibilidades Somáticas - I. Organización General, Las Sensaciones Táctil y Posicional PDFrodrigo4590100% (1)
- BiofluidosDocumento6 pagineBiofluidosAndresNessuna valutazione finora
- Potencial de Acción - FisiologíaDocumento6 paginePotencial de Acción - FisiologíaDara DosNessuna valutazione finora
- Canales IónicosDocumento12 pagineCanales IónicosJulinho M LeOnNessuna valutazione finora
- 20 LaringitisDocumento9 pagine20 LaringitisLilo VotesNessuna valutazione finora
- HipotalamoDocumento4 pagineHipotalamoIrvin Mtz AristaNessuna valutazione finora
- Tejido Epitelial Usmp HistologiaDocumento63 pagineTejido Epitelial Usmp Histologiavibescash100% (1)
- Enfoques PsicologicosDocumento19 pagineEnfoques PsicologicosholasdasdsNessuna valutazione finora
- SubtalamoDocumento50 pagineSubtalamoMariand F Morales100% (2)
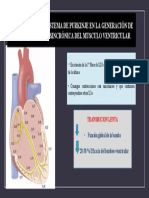
- Importancia Del Sistema de PurkinjeDocumento1 paginaImportancia Del Sistema de PurkinjeCeovi VillanNessuna valutazione finora
- Lipotimia Shock Anafilactico y SincopeDocumento25 pagineLipotimia Shock Anafilactico y SincopeDarwin Mamani ReyesNessuna valutazione finora
- Region Fosa Cigomatica ExpoDocumento14 pagineRegion Fosa Cigomatica ExpoElvis Ponce ColquiNessuna valutazione finora
- Glandula Submandibular.Documento7 pagineGlandula Submandibular.Marcos Erli Banda CubasNessuna valutazione finora
- Nuevo Documento de Microsoft Office WordDocumento5 pagineNuevo Documento de Microsoft Office WordRube Garduño RoaNessuna valutazione finora
- Tarea Sinapsis y Union NeuromuscularDocumento4 pagineTarea Sinapsis y Union NeuromuscularOrlando ComtrerazNessuna valutazione finora
- Señalizacion Celular y Sistema Endocrino PDFDocumento27 pagineSeñalizacion Celular y Sistema Endocrino PDFFiorella Alzamora VignaNessuna valutazione finora
- Anexo de Irrigación y Inervación de ATMDocumento1 paginaAnexo de Irrigación y Inervación de ATMGonzalo San MartinNessuna valutazione finora
- Morfofisiología de La NeuronaDocumento3 pagineMorfofisiología de La Neuronami bella mas bellaNessuna valutazione finora
- Informe Re GolgiDocumento7 pagineInforme Re GolgienriqueNessuna valutazione finora
- Potencial de Accion y MembranaDocumento2 paginePotencial de Accion y MembranaEmily Ortegon100% (1)
- Artículo Científico de MicrosDocumento13 pagineArtículo Científico de MicrosDANIEL ANGEL TORRES ESTUDIANTE ACTIVONessuna valutazione finora
- Cuestionario Tejido NerviosoDocumento6 pagineCuestionario Tejido NerviosoAna BazzanoNessuna valutazione finora
- Termodinámica Aplicada Al HombreDocumento11 pagineTermodinámica Aplicada Al HombreLuis Enrique ArrietaInfanteNessuna valutazione finora
- Vía TrigeminalDocumento11 pagineVía TrigeminalDANIEL ALEJANDRO88% (8)
- 3.ppse y PpsiDocumento63 pagine3.ppse y PpsiJOhn JacoboNessuna valutazione finora
- Resumen de MorfofuncionDocumento8 pagineResumen de MorfofuncionPoly Cab100% (1)
- Historia Natural de La EnfermedadDocumento5 pagineHistoria Natural de La EnfermedadShaid Castañeda YbarrolaNessuna valutazione finora
- Pares Craneales o Nervios CranealesDocumento10 paginePares Craneales o Nervios CranealesAntonio Arroyo CornejoNessuna valutazione finora
- Sentidos EspecialesDocumento8 pagineSentidos EspecialesIon Frias0% (1)
- Anatomia y Fisiologia Del Sistema NerviosoDocumento6 pagineAnatomia y Fisiologia Del Sistema NerviosoCristian AcostaNessuna valutazione finora
- Informe (Cavidad Nasal) - Exposiciòn (Oìdo Medio)Documento21 pagineInforme (Cavidad Nasal) - Exposiciòn (Oìdo Medio)Mary Monja DiazNessuna valutazione finora
- Nervio Vestibulococlear, AnatomíaDocumento3 pagineNervio Vestibulococlear, AnatomíaMartín MeléndezNessuna valutazione finora
- Caso Clinico Clase VDocumento18 pagineCaso Clinico Clase VGisselle CaceresNessuna valutazione finora
- Sistema Nervioso TripticoDocumento2 pagineSistema Nervioso TripticoMariedjalex MarieNessuna valutazione finora
- Neuro Transm I SoresDocumento5 pagineNeuro Transm I Soresdamaris pradoNessuna valutazione finora
- Realizar Un Mapa ConceptualDocumento6 pagineRealizar Un Mapa ConceptualjuanNessuna valutazione finora
- Musculo LisoDocumento9 pagineMusculo LisoMartinVelazcoNessuna valutazione finora
- Cap 48 GuytonDocumento4 pagineCap 48 GuytonCinthia LemusNessuna valutazione finora
- Areas de Brodman de La Corteza HumanaDocumento9 pagineAreas de Brodman de La Corteza HumanaJorge Manuel Pisco ZavaletaNessuna valutazione finora
- Fosa TemporalDocumento18 pagineFosa TemporalJosurer NrtNessuna valutazione finora
- Neuro CompletoDocumento162 pagineNeuro CompletoBrenda PusdáNessuna valutazione finora
- Potencial de Membrana en Reposo de Células ExcitablesDocumento13 paginePotencial de Membrana en Reposo de Células ExcitablesDaniel SantanaNessuna valutazione finora
- Clase Q.F. Alejandra Del Rio ISP Registro Productos Farmaceuticos Biologicos 2021Documento58 pagineClase Q.F. Alejandra Del Rio ISP Registro Productos Farmaceuticos Biologicos 2021ScarlethMuñozNessuna valutazione finora
- Clase 4 Intervalos de ConfianzaDocumento56 pagineClase 4 Intervalos de ConfianzaScarlethMuñozNessuna valutazione finora
- Clase 2Documento48 pagineClase 2ScarlethMuñozNessuna valutazione finora
- Resumen de Técnicas de LaboratorioDocumento20 pagineResumen de Técnicas de LaboratorioScarlethMuñozNessuna valutazione finora
- Humanismo Universidad y CulturaDocumento5 pagineHumanismo Universidad y CulturaEstefania ChangoNessuna valutazione finora
- Topografía I - Teoría de ProbabilidadesDocumento7 pagineTopografía I - Teoría de ProbabilidadesJunior Quispe HuarcayaNessuna valutazione finora
- 1 Generalidades Alcantarillado LRMNDocumento22 pagine1 Generalidades Alcantarillado LRMNLenin MaldonadoNessuna valutazione finora
- Cuadro Comparativo de Las Generaciones de ComputadorasDocumento6 pagineCuadro Comparativo de Las Generaciones de ComputadorasErika Denise Mendez RiosNessuna valutazione finora
- Actividades Tema 13 PDFDocumento4 pagineActividades Tema 13 PDFBruno Bejar AbaldeNessuna valutazione finora
- El Cambio Climático en TacnaDocumento27 pagineEl Cambio Climático en Tacnasupremo7Nessuna valutazione finora
- Presupuesto de Obra - I.E. #523 - Centro Poblado Los AngelesDocumento12 paginePresupuesto de Obra - I.E. #523 - Centro Poblado Los Angelesjose luisNessuna valutazione finora
- CrazyDocumento104 pagineCrazysteffi9112Nessuna valutazione finora
- Términos y Condiciones Mi Cuenta Tigo - 0Documento3 pagineTérminos y Condiciones Mi Cuenta Tigo - 0DS ClanNessuna valutazione finora
- El AdverbioDocumento13 pagineEl AdverbioEsdrasRiosNessuna valutazione finora
- Especificaciones para Camaras Electricas de TransformacionDocumento24 pagineEspecificaciones para Camaras Electricas de TransformacionmanuelNessuna valutazione finora
- Actividad Asociada A CE1.5Documento16 pagineActividad Asociada A CE1.5sofía popNessuna valutazione finora
- Referencia de DispositivosDocumento58 pagineReferencia de Dispositivoscelyn280482Nessuna valutazione finora
- EdMe Strohmaier - Al BiruniDocumento8 pagineEdMe Strohmaier - Al BiruniAngel Chavez EslavaNessuna valutazione finora
- En Que Consiste Cada Maniobra de LiopolDocumento11 pagineEn Que Consiste Cada Maniobra de LiopolAndres BuesoNessuna valutazione finora
- Plan de Clase Semana 7 2016Documento9 paginePlan de Clase Semana 7 2016Cesar MaríaNessuna valutazione finora
- Tarea 3 - Hidrocarburos Aromáticos, Alcoholes y Aminas DIANADELMARDocumento9 pagineTarea 3 - Hidrocarburos Aromáticos, Alcoholes y Aminas DIANADELMARDianaNessuna valutazione finora
- Nom 187 Ssa1 Scfi 2002Documento18 pagineNom 187 Ssa1 Scfi 2002Carolina fuentesNessuna valutazione finora
- Trabajo Construcciones EspecialesDocumento11 pagineTrabajo Construcciones Especialestania reynilda davila vilcaNessuna valutazione finora
- Capitulo Xi - AreasDocumento64 pagineCapitulo Xi - AreasSantos Vasquez AlvitesNessuna valutazione finora
- Libreto de Investigación LavavajillaspdfDocumento41 pagineLibreto de Investigación Lavavajillaspdf7q8dvrr469Nessuna valutazione finora
- WWW - Yoquieroaprobar.es: Fuerzas Y MovimientoDocumento4 pagineWWW - Yoquieroaprobar.es: Fuerzas Y Movimientodjxjud747Nessuna valutazione finora
- Session2 SI 2017Documento23 pagineSession2 SI 2017Vanessa SanchezNessuna valutazione finora
- Formato de Presentación de La ActividadDocumento2 pagineFormato de Presentación de La ActividadJ B100% (2)
- Estrategias y Actividades de MaternalDocumento70 pagineEstrategias y Actividades de MaternalANAPOTTERCULLEN67% (9)
- 16 Proc - Hse - 016 Procedimiento de Autorización de VisitasDocumento11 pagine16 Proc - Hse - 016 Procedimiento de Autorización de VisitasAzumy Lorena Lopez N. AzumyNessuna valutazione finora
- Examen 5 B1 2017-2018Documento18 pagineExamen 5 B1 2017-2018Gustavo GutierrezNessuna valutazione finora
- Hoja de Vida Sendy Yanid CantyDocumento7 pagineHoja de Vida Sendy Yanid CantyKAROLL VIVIANA SÁNCHEZNessuna valutazione finora
- U.ft.12.010.069 Certificacion Determinacion Cedular Ley 1943 de 2018 V1 - FirmadoDocumento1 paginaU.ft.12.010.069 Certificacion Determinacion Cedular Ley 1943 de 2018 V1 - FirmadoКарлос ЦеспедесNessuna valutazione finora