Potrebbero piacerti anche
- 2 libros en 1 - Come para no enfermar: Dime qué comes y te diré de qué vas a enfermar + Probióticos para principiantesDa Everand2 libros en 1 - Come para no enfermar: Dime qué comes y te diré de qué vas a enfermar + Probióticos para principiantesNessuna valutazione finora
- Turbo PascalDocumento353 pagineTurbo PascalCristian Camarena100% (1)
- Edad GestacionalDocumento30 pagineEdad GestacionalAlexanderVillacrésNessuna valutazione finora
- Síndrome del intestino permeable para principiantes - El libro de autoayuda - Cómo interpretar los síntomas del intestino permeable, reconocer las causas y curar tu intestinoDa EverandSíndrome del intestino permeable para principiantes - El libro de autoayuda - Cómo interpretar los síntomas del intestino permeable, reconocer las causas y curar tu intestinoNessuna valutazione finora
- Curso Tecnicas Mantenimiento Control Contaminacion Ajustes Tolerancias Montaje Cojinetes Friccion Desgaste LubricacionDocumento31 pagineCurso Tecnicas Mantenimiento Control Contaminacion Ajustes Tolerancias Montaje Cojinetes Friccion Desgaste LubricacionEliseoNessuna valutazione finora
- Programacion IDocumento166 pagineProgramacion IFrankNessuna valutazione finora
- Manual Borland C++ Builder Con SQL (Espa - Ol)Documento795 pagineManual Borland C++ Builder Con SQL (Espa - Ol)Uriel Alexander LoriaNessuna valutazione finora
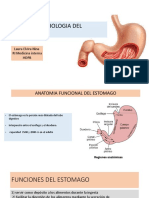
- EstomagoDocumento21 pagineEstomagoLaura Elvira NinaNessuna valutazione finora
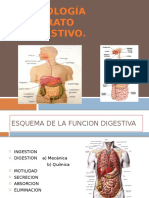
- Fisiologia Del Aparato DigestivoDocumento34 pagineFisiologia Del Aparato DigestivoIlianPlacencia100% (1)
- Practica 3 LlinealidadDocumento10 paginePractica 3 LlinealidadGutslutNessuna valutazione finora
- Festo Fundamentos de ElectroneumaticaDocumento89 pagineFesto Fundamentos de ElectroneumaticaDarío Ordóñez78% (9)
- Cálculo Diferencial Con Matlab - Alejandro Vera LazaroDocumento321 pagineCálculo Diferencial Con Matlab - Alejandro Vera LazaroAlex Galicia67% (3)
- Farmacologia Digestiva PDFDocumento6 pagineFarmacologia Digestiva PDFGianella Michelle Cervantes Del AguilaNessuna valutazione finora
- Castro Jesus Jeronimo - Geometria en Olimpiadas Matematicas PDFDocumento148 pagineCastro Jesus Jeronimo - Geometria en Olimpiadas Matematicas PDFJosé Javier Cruz TorresNessuna valutazione finora
- Trigonometriaq Plana Esferica - Granville, Smith-1Documento315 pagineTrigonometriaq Plana Esferica - Granville, Smith-1N Aguilar AguellesNessuna valutazione finora
- Sopa de Letras SistemasDocumento3 pagineSopa de Letras Sistemasnelfrancisco2Nessuna valutazione finora
- GASTROENTERITISDocumento13 pagineGASTROENTERITISPia LenguaNessuna valutazione finora
- JuanitaDocumento11 pagineJuanitatapiatrujilloximenaNessuna valutazione finora
- Funciones Del EstómagoDocumento3 pagineFunciones Del EstómagoJennifer OyacatoNessuna valutazione finora
- Discusión 3 FarmacoDocumento34 pagineDiscusión 3 FarmacoFátima Guadalupe Guevara BarahonaNessuna valutazione finora
- EnsayoDocumento9 pagineEnsayoMarlene Mercedes Arévalo LinaresNessuna valutazione finora
- Sistema Digestivo GeneralDocumento6 pagineSistema Digestivo GeneraltatiNessuna valutazione finora
- GASTROINTESTINALDocumento21 pagineGASTROINTESTINALManuel Francisco Morales HernándezNessuna valutazione finora
- Secrecion Gástrica y EsofágicaDocumento5 pagineSecrecion Gástrica y EsofágicaJamilex HurtadoNessuna valutazione finora
- Fisiologia Del Estomago 3Documento4 pagineFisiologia Del Estomago 3Zeth MatNessuna valutazione finora
- Tema 18Documento5 pagineTema 18Pablo GarciaNessuna valutazione finora
- Guía 21 RespuestasDocumento3 pagineGuía 21 Respuestasramirezmisa2612Nessuna valutazione finora
- Función Del EstomagoDocumento6 pagineFunción Del EstomagoMaria Jose Perez BarriosNessuna valutazione finora
- Fisio-Digestivo 2016Documento16 pagineFisio-Digestivo 2016Victoria PinascoNessuna valutazione finora
- Funciones Secretoras Del Aparato DigestivoDocumento43 pagineFunciones Secretoras Del Aparato DigestivoJuan Ernesto Gutierrez Sevilla100% (6)
- RP7 VaciamientoDocumento5 pagineRP7 VaciamientoraymundoNessuna valutazione finora
- Antiulcerosos y Antidiarreicos CompletDocumento44 pagineAntiulcerosos y Antidiarreicos Completflor tellez100% (3)
- Resumen Degradando Los AlimentosDocumento7 pagineResumen Degradando Los AlimentosJa NetworkNessuna valutazione finora
- Control de La Secrecion PancreaticaDocumento8 pagineControl de La Secrecion PancreaticaSoany M. Alanoca CoaquiraNessuna valutazione finora
- Funcion Secretora Del Tubo DigestivoDocumento31 pagineFuncion Secretora Del Tubo DigestivoPaola Andrea CANessuna valutazione finora
- Resumen-Fisiología de La Secreción PancreáticaDocumento3 pagineResumen-Fisiología de La Secreción PancreáticaJose Alfredo HernandezNessuna valutazione finora
- Sistema Digestivo. Trabajo FinalDocumento12 pagineSistema Digestivo. Trabajo FinalNayarelis BenaventeNessuna valutazione finora
- Cuestionario Introduccion A La AlimentacionDocumento3 pagineCuestionario Introduccion A La AlimentacionSHIRLEY CUSICANQUINessuna valutazione finora
- Fisiología de La Secreción GástricaDocumento7 pagineFisiología de La Secreción GástricaDamaris ArenasNessuna valutazione finora
- Capitulo 62 GuytonDocumento18 pagineCapitulo 62 GuytonGuadalupe Arenas Rosas100% (1)
- 1 - Regulación de Las Secreciones Del Tracto DigestivoDocumento60 pagine1 - Regulación de Las Secreciones Del Tracto DigestivoAndrea MozoNessuna valutazione finora
- Fisiologìa Práctica - Informe 14 (Rcruzp-6)Documento6 pagineFisiologìa Práctica - Informe 14 (Rcruzp-6)Vanessa AvilaNessuna valutazione finora
- Guia de Fisiologia de La Secrecion GastricaDocumento9 pagineGuia de Fisiologia de La Secrecion GastricaIsaNessuna valutazione finora
- Enfe Unidad 6 (7) Digestivo 2023 PDFDocumento24 pagineEnfe Unidad 6 (7) Digestivo 2023 PDFJONATAN LUIS LEDESMANessuna valutazione finora
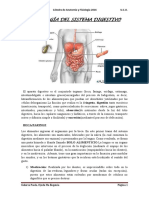
- Httpsaulavirtual - Uninorte.edu - Pypluginfile.phpfile 2F61885092Fmod resource2Fcontent2F02FAparato20Digestivo20-20PresDocumento22 pagineHttpsaulavirtual - Uninorte.edu - Pypluginfile.phpfile 2F61885092Fmod resource2Fcontent2F02FAparato20Digestivo20-20PresPablo CenturiónNessuna valutazione finora
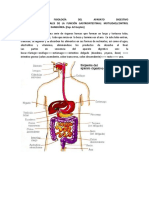
- Aparato DigestivoDocumento50 pagineAparato DigestivoTammy GonzalezNessuna valutazione finora
- Bioquimica-Clase 5Documento30 pagineBioquimica-Clase 5Ana LópezNessuna valutazione finora
- Sem 1 Monografia de La Secrecion Del Tracto GastrointestinalDocumento11 pagineSem 1 Monografia de La Secrecion Del Tracto GastrointestinalBrayan CaipoNessuna valutazione finora
- Funciones Digestivas 2020Documento45 pagineFunciones Digestivas 2020jeniffer Ym100% (1)
- Patologia IiDocumento19 paginePatologia IiYosajandy JimenezNessuna valutazione finora
- Casos Clinicos y PreguntasDocumento4 pagineCasos Clinicos y PreguntasAngheles FrontanillaNessuna valutazione finora
- DigestiónDocumento10 pagineDigestiónAlan PonceNessuna valutazione finora
- Antiulcerosos y Antidiarreicos CompletDocumento42 pagineAntiulcerosos y Antidiarreicos Completflor tellezNessuna valutazione finora
- Fisiologia Nutricion Clase 10 y 11Documento60 pagineFisiologia Nutricion Clase 10 y 11NAYELI LORENA HUARZAYA CUPINessuna valutazione finora
- Funciones Secretoras Del Tubo Digestivo Fisio2017Documento5 pagineFunciones Secretoras Del Tubo Digestivo Fisio2017Karla DuarteNessuna valutazione finora
- SecretinaDocumento6 pagineSecretinaHermán GeronimoNessuna valutazione finora
- ExpoDocumento6 pagineExpoYOVANNY ALEXANDER BENAVIDES VALLEJONessuna valutazione finora
- Fisiologia Del EstomagoDocumento4 pagineFisiologia Del EstomagoPau BautistaNessuna valutazione finora
- GastrointestinalDocumento25 pagineGastrointestinalCatalina Cané Muñoz SilvaNessuna valutazione finora
- Sistema DigestivoDocumento30 pagineSistema DigestivoBriseida Yareth Ibarra ZapataNessuna valutazione finora
- Fase Cefalica HCL PDFDocumento5 pagineFase Cefalica HCL PDFadalithNessuna valutazione finora
- Secrecion GastricaDocumento2 pagineSecrecion GastricaChiquinquira GarciaNessuna valutazione finora
- Fisiologia Del Aparato Digestivo 2014 MDocumento46 pagineFisiologia Del Aparato Digestivo 2014 MSilvana Ruelas EliasNessuna valutazione finora
- Pancreas NOTAS MEDICASDocumento2 paginePancreas NOTAS MEDICASLuis ValdezNessuna valutazione finora
- Secreciones DigestivasDocumento5 pagineSecreciones DigestivasIrving CastilloNessuna valutazione finora
- Funciones Secretoras Del Aparato DigestivoDocumento73 pagineFunciones Secretoras Del Aparato DigestivoJosé DaNessuna valutazione finora
- Células Mucosas Superficiales - EstomagoDocumento2 pagineCélulas Mucosas Superficiales - EstomagoTatiana RamirezNessuna valutazione finora
- Ulcera Peptica - Fisiopatología PDFDocumento39 pagineUlcera Peptica - Fisiopatología PDFAnd1f2 RNessuna valutazione finora
- Tema 16Documento10 pagineTema 16baselsheikh2022Nessuna valutazione finora
- Tipos DatosDocumento2 pagineTipos DatosNixon RondonNessuna valutazione finora
- Manual MacrosDocumento44 pagineManual MacrosVanessa KarinaNessuna valutazione finora
- Programación CDocumento57 pagineProgramación Cgraduadoesime50% (2)
- Metodo de Secante y Regula Falsi-2Documento6 pagineMetodo de Secante y Regula Falsi-2AlexanderVillacrésNessuna valutazione finora
- Universidad Nacional de ChimborazoDocumento1 paginaUniversidad Nacional de ChimborazoAlexanderVillacrésNessuna valutazione finora
- Universidad Nacional de ChimborazoDocumento1 paginaUniversidad Nacional de ChimborazoAlexanderVillacrésNessuna valutazione finora
- InterpolacionDocumento4 pagineInterpolacionEl EDu Arrow RodasNessuna valutazione finora
- Reaccionesendisolucinacuosa 160728215835 PDFDocumento32 pagineReaccionesendisolucinacuosa 160728215835 PDFAlexanderVillacrésNessuna valutazione finora
- 04 Reacciones en Soluicion AcuosaDocumento32 pagine04 Reacciones en Soluicion AcuosaefuenzalidaqNessuna valutazione finora
- URTICARIADocumento1 paginaURTICARIAAlexanderVillacrésNessuna valutazione finora
- Enfermedad de AddisonDocumento13 pagineEnfermedad de AddisonAlexanderVillacrésNessuna valutazione finora
- Reacciones QuimicasDocumento7 pagineReacciones QuimicasAlexanderVillacrésNessuna valutazione finora
- Reacciones QuimicasDocumento7 pagineReacciones QuimicasAlexanderVillacrésNessuna valutazione finora
- HIDROLISISDocumento11 pagineHIDROLISISDanny QuilcaNessuna valutazione finora
- Enfermedad de AddisonDocumento13 pagineEnfermedad de AddisonAlexanderVillacrésNessuna valutazione finora
- UrticariaDocumento15 pagineUrticariaAlexanderVillacrésNessuna valutazione finora
- Cetoacidosis DiabéticaDocumento16 pagineCetoacidosis DiabéticaAlexanderVillacrésNessuna valutazione finora
- Edad GestacionalDocumento30 pagineEdad GestacionalAlexanderVillacrésNessuna valutazione finora
- Compuestos OxigenadosDocumento13 pagineCompuestos OxigenadosAlexanderVillacrésNessuna valutazione finora
- Poli Trauma Tiz A DoDocumento5 paginePoli Trauma Tiz A DoAlexanderVillacrésNessuna valutazione finora
- Efectos SecundariosDocumento2 pagineEfectos SecundariosAlexanderVillacrésNessuna valutazione finora
- Clase 05 Curso 3MDocumento5 pagineClase 05 Curso 3Mdaniel diazNessuna valutazione finora
- EcocardiogramaDocumento23 pagineEcocardiogramaSandy CyndyNessuna valutazione finora
- 8 INS Balanza Dibal SERIE LDocumento159 pagine8 INS Balanza Dibal SERIE LJosué Gonzaga71% (7)
- Práctica N 4 FINALDocumento20 paginePráctica N 4 FINALEmerson Casi MarroNessuna valutazione finora
- Dada La Siguiente TopologíaDocumento7 pagineDada La Siguiente Topologíajaulloa1Nessuna valutazione finora
- La Oración Es La Unidad de Expresión en El IdiomaDocumento6 pagineLa Oración Es La Unidad de Expresión en El IdiomaMoasibeaNessuna valutazione finora
- TrasductorDocumento6 pagineTrasductorjohnny Ivan Choque lopezNessuna valutazione finora
- PH en El Agua TripticoDocumento2 paginePH en El Agua TripticoAndyHerranGilesNessuna valutazione finora
- Estructuras HiperestaticasDocumento12 pagineEstructuras Hiperestaticasfranklin jimenez mendezNessuna valutazione finora
- Tarea 1: Concepto IntegralDocumento48 pagineTarea 1: Concepto Integralsteven baronNessuna valutazione finora
- Equilibrio Químico. KC y KP: 1.-La Constante de Equilibrio KDocumento2 pagineEquilibrio Químico. KC y KP: 1.-La Constante de Equilibrio KLuis Mario 607 Vasquez RomeroNessuna valutazione finora
- Sistema PortacablesDocumento8 pagineSistema PortacablesChristian Rhadames Barkley ChavezNessuna valutazione finora
- Requerimientos Conservas Carnicas2018Documento34 pagineRequerimientos Conservas Carnicas2018QUINTOSLLANOSNessuna valutazione finora
- 5to y 6yo TALENTOSDocumento6 pagine5to y 6yo TALENTOSguillermo henrryNessuna valutazione finora
- Presentacion OdorizadorDocumento25 paginePresentacion OdorizadorMiguel Jiménez FloresNessuna valutazione finora
- Taller 1 - Cálculo ProposicionalDocumento11 pagineTaller 1 - Cálculo ProposicionalCesar VillanuevaNessuna valutazione finora
- EbullicionDocumento8 pagineEbullicionMonsif AhlikNessuna valutazione finora
- Pruebas en La LechadaDocumento5 paginePruebas en La LechadaJairo MataNessuna valutazione finora
- Parcial de Areas y PerimetrosDocumento4 pagineParcial de Areas y Perimetrosbibiana farley vera yara100% (1)
- COLOIDEDocumento2 pagineCOLOIDEWilber Mamáni PerezNessuna valutazione finora
- La Forma Correcta de Lubricar Engranajes Corona-SinfínDocumento7 pagineLa Forma Correcta de Lubricar Engranajes Corona-SinfínHarold none03Nessuna valutazione finora
- Rutas de Recolección SIGDocumento25 pagineRutas de Recolección SIGJuan Antonio Araiza AguilarNessuna valutazione finora
- Matemáticas para Los Negocios - Semana 3 - PDocumento8 pagineMatemáticas para Los Negocios - Semana 3 - PBelén IbarraNessuna valutazione finora
- 1 InfografiasDocumento4 pagine1 InfografiasTatiiana BohórquezNessuna valutazione finora
- Propiedades Del Gas Natural y El Factor de Compresibilidad Del GasDocumento18 paginePropiedades Del Gas Natural y El Factor de Compresibilidad Del GasMarcos Daniel Vargas InocheaNessuna valutazione finora
- Soluciones QuimicasDocumento48 pagineSoluciones QuimicasDaysi Taborda RamírezNessuna valutazione finora