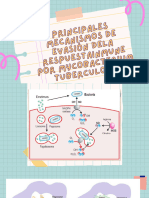
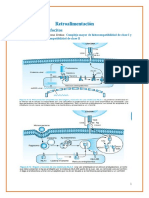
Potrebbero piacerti anche
- Fundamentos y técnicas básicas en biología molecularDa EverandFundamentos y técnicas básicas en biología molecularNessuna valutazione finora
- Transferencia de inmunidad en el binomio madre-cría: Estudio comparativo entre especiesDa EverandTransferencia de inmunidad en el binomio madre-cría: Estudio comparativo entre especiesNessuna valutazione finora
- 03 Que Son Los InflamasomasDocumento8 pagine03 Que Son Los InflamasomasLeon-lv Londoño100% (1)
- Receptores Citosólicos para Pamp y DampDocumento4 pagineReceptores Citosólicos para Pamp y DampPame FloresNessuna valutazione finora
- Seminario 2. Inmunidad Innata. Parte 2.Documento29 pagineSeminario 2. Inmunidad Innata. Parte 2.lilianabarbagallo.estilistaNessuna valutazione finora
- Reseñas: Inflamasomas: Mecanismo de Ensamblaje, Regulación y SeñalizaciónDocumento14 pagineReseñas: Inflamasomas: Mecanismo de Ensamblaje, Regulación y Señalizacióntenma1200Nessuna valutazione finora
- Fagocitosis y PRRDocumento4 pagineFagocitosis y PRRisrael tulcanazaNessuna valutazione finora
- El Papel de Los Receptores de Reconocimiento de Patrones en La Inmunidad InnataDocumento46 pagineEl Papel de Los Receptores de Reconocimiento de Patrones en La Inmunidad InnataKArdo23Nessuna valutazione finora
- La Enfermedad Del Sueño Tiene Sus Orígenes en La Región Afrotropical Del ContinenteDocumento4 pagineLa Enfermedad Del Sueño Tiene Sus Orígenes en La Región Afrotropical Del ContinentejordanNessuna valutazione finora
- Receptores CitosolicosDocumento33 pagineReceptores CitosolicosVanessa YapoNessuna valutazione finora
- Receptores SolublesDocumento6 pagineReceptores SolublesCarlitos AyalaNessuna valutazione finora
- PRRsDocumento3 paginePRRsOmar Adrian Hernandez JimenezNessuna valutazione finora
- Taller Sobre CitoesqueletoDocumento9 pagineTaller Sobre CitoesqueletoNatalie Rodriguez CortésNessuna valutazione finora
- Innata 2Documento46 pagineInnata 2henriquecastelanNessuna valutazione finora
- Exposición GrupalDocumento27 pagineExposición GrupalStephania SorianoNessuna valutazione finora
- Capítulo I - Respuesta Inmune. de La Fisiología A La MedicinaDocumento26 pagineCapítulo I - Respuesta Inmune. de La Fisiología A La Medicinaicaro chaineNessuna valutazione finora
- Papel de Los Receptores Tipo Toll en La Inmunidad InnataDocumento6 paginePapel de Los Receptores Tipo Toll en La Inmunidad InnataG-orgina Coello0% (1)
- Resumen - Generalidades Del Reconocimiento Por La Inmunidad InnataDocumento7 pagineResumen - Generalidades Del Reconocimiento Por La Inmunidad InnataRanciovichNessuna valutazione finora
- Inmunidad InnataDocumento22 pagineInmunidad InnataIsabellaNessuna valutazione finora
- Patologia y CausaDocumento7 paginePatologia y CausaAndrea K. CuelloNessuna valutazione finora
- Proceso de TranscripciónDocumento17 pagineProceso de TranscripciónKarina Alejandra Cervantes RoblesNessuna valutazione finora
- T4 InmunoDocumento21 pagineT4 InmunoCris SánchezNessuna valutazione finora
- 3 - PAMPs y RRPDocumento3 pagine3 - PAMPs y RRPGino MartinezNessuna valutazione finora
- Investigaciones InmunologiaDocumento15 pagineInvestigaciones InmunologiaErick Salazar ManzanoNessuna valutazione finora
- Actividad Receptores de PRRDocumento11 pagineActividad Receptores de PRRAndres PelayoNessuna valutazione finora
- Marco TeoricoDocumento46 pagineMarco TeoricoLiam RodriguezNessuna valutazione finora
- Presentacion Receptores Citosolicos y de MembranaDocumento41 paginePresentacion Receptores Citosolicos y de Membranaisabella vNessuna valutazione finora
- Respuesta InnataDocumento4 pagineRespuesta InnataAndrea Mariel PeñaNessuna valutazione finora
- Receptores Tipo Toll - TareasDocumento12 pagineReceptores Tipo Toll - TareasDayana Torrico VNessuna valutazione finora
- Receptores Tipo Toll - TareasDocumento12 pagineReceptores Tipo Toll - TareasDayana Torrico VNessuna valutazione finora
- Receptores Tipo Toll (TLR)Documento36 pagineReceptores Tipo Toll (TLR)Brenda Collantes VidalNessuna valutazione finora
- Paramyxoviridae TEXTODocumento28 pagineParamyxoviridae TEXTOSharon ReyesNessuna valutazione finora
- El Papel de Los TLR en La Activación de NeutrófilosDocumento6 pagineEl Papel de Los TLR en La Activación de NeutrófilosVic VargasNessuna valutazione finora
- Inflamación y Enfermedad de ParkinsonDocumento19 pagineInflamación y Enfermedad de Parkinsonjuan carlos muñozNessuna valutazione finora
- Receptores Del Tipo RIGDocumento7 pagineReceptores Del Tipo RIGReyner CovarrubiasNessuna valutazione finora
- Retroalimentación Area 2Documento22 pagineRetroalimentación Area 2Dove CameronNessuna valutazione finora
- Fichas 7Documento7 pagineFichas 7Noelita RiveraNessuna valutazione finora
- Papel Regulador Del ATPDocumento7 paginePapel Regulador Del ATPAlejandro DelgadoNessuna valutazione finora
- 169 Sistema Inmunitario Cutáneo PDFDocumento63 pagine169 Sistema Inmunitario Cutáneo PDFJ C Torres Formalab100% (1)
- LPS y Receptores Toll (MAOG)Documento13 pagineLPS y Receptores Toll (MAOG)miguel angel100% (3)
- 2do Parcial Medicina MolecularDocumento7 pagine2do Parcial Medicina MolecularAna Daniela Cruz ZarabiaNessuna valutazione finora
- Respuesta Inmune Innata - 1ra ParteDocumento18 pagineRespuesta Inmune Innata - 1ra Parteirving hilarioNessuna valutazione finora
- Mecanismos de Transcripcion.Documento41 pagineMecanismos de Transcripcion.Jhoe100% (1)
- 5 Simposios 105-340Documento89 pagine5 Simposios 105-340EDUARDO CARRILLO GODINEZNessuna valutazione finora
- HETERÓFILOSDocumento3 pagineHETERÓFILOSRosa Mezta DamisNessuna valutazione finora
- NEUROTROFINASDocumento6 pagineNEUROTROFINASSandra chopinNessuna valutazione finora
- 4.moleculas Del Sistema Inmunologico EspecificoDocumento66 pagine4.moleculas Del Sistema Inmunologico EspecificoGonzales Sanchez JoseNessuna valutazione finora
- Sepsis MaternaDocumento40 pagineSepsis MaternaAlberto BrunoNessuna valutazione finora
- Resumen - Clase Teórica de Virus Herpes Simplex y Varicela-ZósterDocumento4 pagineResumen - Clase Teórica de Virus Herpes Simplex y Varicela-Zósterapi-3697245100% (3)
- Receptores NKDocumento2 pagineReceptores NKToledo YamilaNessuna valutazione finora
- Inmunidad InnataDocumento14 pagineInmunidad InnataMelissa RodriguezNessuna valutazione finora
- Alonso, Neuropéptidos, Estrés y AlergiaDocumento16 pagineAlonso, Neuropéptidos, Estrés y AlergiaPC Marian RjNessuna valutazione finora
- Tarea S10 Histología MédicaDocumento8 pagineTarea S10 Histología MédicaNikola García RiveraNessuna valutazione finora
- Tema 5. Reconocimiento en La Inmunidad InnataDocumento5 pagineTema 5. Reconocimiento en La Inmunidad InnatalucisNessuna valutazione finora
- Sem1 InmunoDocumento79 pagineSem1 InmunoRicardo DanielNessuna valutazione finora
- El Proceso de Expresión Génica2Documento21 pagineEl Proceso de Expresión Génica2Erika PerezNessuna valutazione finora
- 572-Texto Del Artículo-1074-1-10-20190702 PDFDocumento10 pagine572-Texto Del Artículo-1074-1-10-20190702 PDFFernanda AltamiranoNessuna valutazione finora
- Inmunidad InnataDocumento38 pagineInmunidad InnataAnyello BuitragoNessuna valutazione finora
- NomenclaturaDocumento3 pagineNomenclaturaNadia Araya AvendañoNessuna valutazione finora
- La Genética en La ActualidadDocumento28 pagineLa Genética en La Actualidadeddy leonNessuna valutazione finora
- Consenso Psoriasis. Actualizacion 2019 14112019Documento22 pagineConsenso Psoriasis. Actualizacion 2019 14112019Giorjhian Erick Lomas ArenasNessuna valutazione finora
- Gui As Basadas en La Evidencia para El Manejo de La Psoriasis en Colombia - EsDocumento257 pagineGui As Basadas en La Evidencia para El Manejo de La Psoriasis en Colombia - Esdarturo67Nessuna valutazione finora
- Mehu100 U1 t9 Sindromes Mielodisplasicos GoldmanDocumento7 pagineMehu100 U1 t9 Sindromes Mielodisplasicos GoldmanMarijose Salas VargasNessuna valutazione finora
- Mehu100 - U1 - t9 - Sindromes Mielodisplasicos y Neoplasias RozmannDocumento5 pagineMehu100 - U1 - t9 - Sindromes Mielodisplasicos y Neoplasias RozmannMarijose Salas VargasNessuna valutazione finora
- Mehu100 U1 t9 Sindromes Mielodisplasicos GoldmanDocumento7 pagineMehu100 U1 t9 Sindromes Mielodisplasicos GoldmanMarijose Salas VargasNessuna valutazione finora
- Guias Clinica Psoariasis ChileDocumento19 pagineGuias Clinica Psoariasis ChileCarlos DNessuna valutazione finora
- 2.proteccion RadiologicaDocumento8 pagine2.proteccion RadiologicaMarijose Salas VargasNessuna valutazione finora
- Resultados y Discusión de ResultadosDocumento6 pagineResultados y Discusión de ResultadosMarijose Salas VargasNessuna valutazione finora
- Fisiología Tiroides y ParatiroidesDocumento43 pagineFisiología Tiroides y ParatiroidesMarijose Salas VargasNessuna valutazione finora
- Lectura Previa 2 - HCL ElectrónicaDocumento22 pagineLectura Previa 2 - HCL ElectrónicaMarijose Salas VargasNessuna valutazione finora
- Silabo - ParasitologíaDocumento12 pagineSilabo - ParasitologíaMarijose Salas Vargas100% (2)
- Marco TeoricoDocumento3 pagineMarco TeoricoMarijose Salas VargasNessuna valutazione finora
- Farmaco Informe IiiDocumento21 pagineFarmaco Informe IiiMarijose Salas VargasNessuna valutazione finora
- Tiroides - Doctor SalazarDocumento78 pagineTiroides - Doctor SalazarMarijose Salas VargasNessuna valutazione finora
- Clase Medula p2Documento44 pagineClase Medula p2Marijose Salas VargasNessuna valutazione finora
- Practicas de Lab. Morfofisiologia PDFDocumento91 paginePracticas de Lab. Morfofisiologia PDFMarijose Salas VargasNessuna valutazione finora
- Citogenética Humana Oncohematologia PDFDocumento8 pagineCitogenética Humana Oncohematologia PDFjhonandyramosNessuna valutazione finora
- Inmunidad Adaptativa y Presentacion de AntigenoDocumento34 pagineInmunidad Adaptativa y Presentacion de AntigenoAna Flores CahuayaNessuna valutazione finora
- Reacción en Cadena de La Polimerasa (PCR)Documento20 pagineReacción en Cadena de La Polimerasa (PCR)Jhon Kevin Bonilla LeónNessuna valutazione finora
- Manual VIHDocumento36 pagineManual VIHMemo TutNessuna valutazione finora
- Tema 1 Introduccion A La Inmunologia 2020-2021 x2Documento17 pagineTema 1 Introduccion A La Inmunologia 2020-2021 x2León Felipe Ureña100% (1)
- Biologia Euclides 01Documento14 pagineBiologia Euclides 01stiven guerrero paivaNessuna valutazione finora
- Mapa Conceptual Sistema InmuneDocumento2 pagineMapa Conceptual Sistema InmunedenisseNessuna valutazione finora
- Laringitis Aguda y CrónicaDocumento14 pagineLaringitis Aguda y CrónicaCarly Arratia Valdez50% (2)
- Enter o CoccusDocumento47 pagineEnter o Coccusbilly casasNessuna valutazione finora
- Promsa InformesDocumento2 paginePromsa InformesZerjmaster19Nessuna valutazione finora
- Clase #5 Genoma HumanoDocumento34 pagineClase #5 Genoma HumanoAsafh FernandezNessuna valutazione finora
- Tema 1Documento30 pagineTema 1Paula EscobarNessuna valutazione finora
- EnterobacteriasDocumento13 pagineEnterobacteriasAbigail VCNessuna valutazione finora
- FINAL DE BIOLOGÍA - 1RA MARZO 2022-MescladoDocumento67 pagineFINAL DE BIOLOGÍA - 1RA MARZO 2022-MescladoTercio SANTOSNessuna valutazione finora
- Lectura 3aDocumento16 pagineLectura 3aYerlin Huamán BautistaNessuna valutazione finora
- Guia7-Sistema-Inmune en Los Seres VivosDocumento18 pagineGuia7-Sistema-Inmune en Los Seres VivosMaria Elena SaraviaNessuna valutazione finora
- Glosario de Anatomia Patologica (4ta Semana) .Documento6 pagineGlosario de Anatomia Patologica (4ta Semana) .JUAN FRANCISCO GOMEZ CERINONessuna valutazione finora
- Generalidades de Los AntibióticosDocumento11 pagineGeneralidades de Los AntibióticosDiego SalinasNessuna valutazione finora
- Modulo 1 Asistente de Laboratorio y ExtraccionistaDocumento212 pagineModulo 1 Asistente de Laboratorio y Extraccionistaveradebora602Nessuna valutazione finora
- Fito 9Documento17 pagineFito 9Jose Alcantara VergaraNessuna valutazione finora
- MENINGITISDocumento49 pagineMENINGITISDiana Janeth García ZamoraNessuna valutazione finora
- ApicomplexaDocumento42 pagineApicomplexaPedro PalmaNessuna valutazione finora
- EmbDocumento3 pagineEmbana mariaNessuna valutazione finora
- Cuadro SinopticoDocumento1 paginaCuadro SinopticoPepe Mejia100% (2)
- Antibioticos y Quimioterapeuticos, Resistencia, Mecanismos de AcciónDocumento90 pagineAntibioticos y Quimioterapeuticos, Resistencia, Mecanismos de AcciónJosé Manuel100% (16)
- CuestionarioDocumento4 pagineCuestionarioRogelioNessuna valutazione finora
- Guías Prácticas de GenéticaDocumento27 pagineGuías Prácticas de GenéticaLuis SierraNessuna valutazione finora
- Apuntes Pruebas BioqimicasDocumento40 pagineApuntes Pruebas BioqimicasAbii De OsorioNessuna valutazione finora
- Evaluacion Final Periodo Iv Noveno 2015Documento2 pagineEvaluacion Final Periodo Iv Noveno 2015Lina Luz Perez MedranoNessuna valutazione finora