Potrebbero piacerti anche
- Total Scar Management: From Lasers to Surgery for Scars, Keloids, and Scar ContracturesDa EverandTotal Scar Management: From Lasers to Surgery for Scars, Keloids, and Scar ContracturesRei OgawaNessuna valutazione finora
- Stem Cells and Cell Signalling in Skeletal MyogenesisDa EverandStem Cells and Cell Signalling in Skeletal MyogenesisNessuna valutazione finora
- Skeletal Muscle Hypertrophy and Regeneration Interplay Between The Myogenic Regulatory Factors (MRFS) and Insulin Like Growth Factors (IGFs) PathwaysDocumento14 pagineSkeletal Muscle Hypertrophy and Regeneration Interplay Between The Myogenic Regulatory Factors (MRFS) and Insulin Like Growth Factors (IGFs) PathwaysGuizzudo CraftNessuna valutazione finora
- Therapeutic Effect of Passive Mobilization Exercise On Improvement of Muscle Regeneration and Prevention of Fibrosis After Laceration Injury of RatDocumento7 pagineTherapeutic Effect of Passive Mobilization Exercise On Improvement of Muscle Regeneration and Prevention of Fibrosis After Laceration Injury of RatCristhian MellaNessuna valutazione finora
- 2023 Article 573Documento15 pagine2023 Article 573orellanaroco.10Nessuna valutazione finora
- Cellular and Molecular Pathways Controlling Muscle Size in Response To ExerciseDocumento29 pagineCellular and Molecular Pathways Controlling Muscle Size in Response To ExerciseMarija NikolicNessuna valutazione finora
- Nature, 2023 - Exercise Metabolism and Adaptation in Skeletal MuscleDocumento26 pagineNature, 2023 - Exercise Metabolism and Adaptation in Skeletal MuscleAlmir FilsNessuna valutazione finora
- Jphysiol 2011 221200Documento12 pagineJphysiol 2011 221200Donato FormicolaNessuna valutazione finora
- The Influence of Anti-Inflammatory Medication On Exercise-Induced Myogenic Precursor Cell Responses in HumansDocumento8 pagineThe Influence of Anti-Inflammatory Medication On Exercise-Induced Myogenic Precursor Cell Responses in Humanssaulo_martorelliNessuna valutazione finora
- Mackey Et Al 2007 The Influence of Anti Inflammatory Medication On Exercise Induced Myogenic Precursor Cell ResponsesDocumento7 pagineMackey Et Al 2007 The Influence of Anti Inflammatory Medication On Exercise Induced Myogenic Precursor Cell Responsesgung26598Nessuna valutazione finora
- Regeneracion Lesiones 2020Documento28 pagineRegeneracion Lesiones 2020CARLOSNessuna valutazione finora
- 10 1152@japplphysiol 00917 2018Documento35 pagine10 1152@japplphysiol 00917 2018cristianNessuna valutazione finora
- Recovery of The Immune System After ExerciseDocumento11 pagineRecovery of The Immune System After Exercisejoao pedroNessuna valutazione finora
- Effects of Training, Detraining, and Retraining On Strength, Hypertrophy, and Myonuclear Number in Human Skeletal MuscleDocumento10 pagineEffects of Training, Detraining, and Retraining On Strength, Hypertrophy, and Myonuclear Number in Human Skeletal Musclejhonnydm93Nessuna valutazione finora
- BBA - Molecular Cell Research: ReviewDocumento11 pagineBBA - Molecular Cell Research: ReviewiuventasNessuna valutazione finora
- Dosagem Otima Musculo PDFDocumento10 pagineDosagem Otima Musculo PDFJanaina PanizzaNessuna valutazione finora
- Marimuthu, 2011Documento7 pagineMarimuthu, 2011Jor ChavezNessuna valutazione finora
- Denervated Muscle Extract Promotes Recovery of Muscle Atrophy Through Activation of Satellite Cells. An Experimental StudyDocumento9 pagineDenervated Muscle Extract Promotes Recovery of Muscle Atrophy Through Activation of Satellite Cells. An Experimental StudyIvana DjolovicNessuna valutazione finora
- The Kynurenine Connection: How Exercise Shifts Muscle Tryptophan Metabolism and Affects Energy Homeostasis, The Immune System, and The BrainDocumento13 pagineThe Kynurenine Connection: How Exercise Shifts Muscle Tryptophan Metabolism and Affects Energy Homeostasis, The Immune System, and The Brainmarisol hernandezNessuna valutazione finora
- Hicks Et Al 2012 Mechanical Strain Applied To Human Fibroblasts Differentially Regulates Skeletal MyoblastDocumento8 pagineHicks Et Al 2012 Mechanical Strain Applied To Human Fibroblasts Differentially Regulates Skeletal MyoblasthjakamjNessuna valutazione finora
- A Cellular Memory Mechanism Aids Overload Hypertrophy in MuscleDocumento10 pagineA Cellular Memory Mechanism Aids Overload Hypertrophy in MuscleGuilherme ReisNessuna valutazione finora
- Bruusgaard 2010Documento7 pagineBruusgaard 2010Sava DarpiniNessuna valutazione finora
- Review Article: Treatment of Skeletal Muscle Injury: A ReviewDocumento8 pagineReview Article: Treatment of Skeletal Muscle Injury: A ReviewkotraeNessuna valutazione finora
- Kagramian FSHD Report 1Documento16 pagineKagramian FSHD Report 1api-700030358Nessuna valutazione finora
- Ajpregu 00060 2002Documento10 pagineAjpregu 00060 2002Jessi LaurentiusNessuna valutazione finora
- Materi PIR - Dr. Imam Subadi, SP - KFR-K Cellular Adaption in Exercise Movement and Exercise in Pain ManagementDocumento29 pagineMateri PIR - Dr. Imam Subadi, SP - KFR-K Cellular Adaption in Exercise Movement and Exercise in Pain ManagementGhea Putri HendrianiNessuna valutazione finora
- Local Immunomodulation and Muscle Progenitor Cells Induce Recovery in Atrophied Muscles in Spinal Cord Injury PatientsDocumento10 pagineLocal Immunomodulation and Muscle Progenitor Cells Induce Recovery in Atrophied Muscles in Spinal Cord Injury PatientsGustavo MovigliaNessuna valutazione finora
- Recovery of Function in Skeletal Muscle Following 2 Different Contraction-Induced InjuriesDocumento9 pagineRecovery of Function in Skeletal Muscle Following 2 Different Contraction-Induced InjuriesCristhian MellaNessuna valutazione finora
- Fisio y EmociònDocumento12 pagineFisio y EmociònDiana BravoNessuna valutazione finora
- Satellite Cell Numbers in Young and Older Men 24 Hours After Eccentric ExerciseDocumento12 pagineSatellite Cell Numbers in Young and Older Men 24 Hours After Eccentric ExerciseGorkaBuesaNessuna valutazione finora
- Bioengineering 10 00956Documento22 pagineBioengineering 10 00956Pieter van KerkhofNessuna valutazione finora
- Functional Electrical Stimulation of Denervated MusclesDocumento7 pagineFunctional Electrical Stimulation of Denervated MusclesMirko GrausoNessuna valutazione finora
- Ibata & Terentjev 2021 - Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadDocumento15 pagineIbata & Terentjev 2021 - Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadIvan TaranovNessuna valutazione finora
- Skeletal Muscle Regeneration Is Modulated by Inflammation: SciencedirectDocumento8 pagineSkeletal Muscle Regeneration Is Modulated by Inflammation: SciencedirectRendra Syani Ulya FitriNessuna valutazione finora
- An Evidence Based Narrative Review of Mechanisms.16Documento14 pagineAn Evidence Based Narrative Review of Mechanisms.16Philippus Von HohenheimNessuna valutazione finora
- Sleep DeprivationDocumento7 pagineSleep DeprivationMayuri BishtNessuna valutazione finora
- DOMS Indicador de EFECTIVIDADDocumento12 pagineDOMS Indicador de EFECTIVIDADNicolás ManonniNessuna valutazione finora
- Isoinercial Performance 2018Documento12 pagineIsoinercial Performance 2018abcsouzaNessuna valutazione finora
- JCSM 9 631Documento4 pagineJCSM 9 631Nakarit SangsirinawinNessuna valutazione finora
- Role of Satellite Cells in Muscle Growth and Maintenance of Muscle MassDocumento7 pagineRole of Satellite Cells in Muscle Growth and Maintenance of Muscle MassLucas Guimarães FerreiraNessuna valutazione finora
- Effect of Explosive Resistance Training On Titin.4Documento7 pagineEffect of Explosive Resistance Training On Titin.4Gust AvoNessuna valutazione finora
- Exer KinesDocumento9 pagineExer KinesNasir AbbasNessuna valutazione finora
- Xie 2006Documento8 pagineXie 2006Affan Nadeem QaziNessuna valutazione finora
- Uncorrected Author Proof: Molecular Mechanisms of Skeletal Muscle HypertrophyDocumento15 pagineUncorrected Author Proof: Molecular Mechanisms of Skeletal Muscle HypertrophyanjosNessuna valutazione finora
- Aps 171 3 233-239 2001Documento7 pagineAps 171 3 233-239 2001Adriano de AssisNessuna valutazione finora
- Review Article: Exercise Training Promotes Functional Recovery After Spinal Cord InjuryDocumento7 pagineReview Article: Exercise Training Promotes Functional Recovery After Spinal Cord InjuryWahyu Satria KurniawanNessuna valutazione finora
- Does Exercise Increase or Decrease Pain Fin 20-03-19Documento37 pagineDoes Exercise Increase or Decrease Pain Fin 20-03-19arifwahyudiNessuna valutazione finora
- Vitro MPC Pro-Liferation and DifferentiationDocumento14 pagineVitro MPC Pro-Liferation and DifferentiationNev NnNessuna valutazione finora
- Moderate Intensity Physical Training Accelerates Healing of Full-Thickness Wounds in MiceDocumento12 pagineModerate Intensity Physical Training Accelerates Healing of Full-Thickness Wounds in MiceGlauber Souza TRANessuna valutazione finora
- Exercise Training Increases Mitochondrial Biogenesis in The BrainDocumento6 pagineExercise Training Increases Mitochondrial Biogenesis in The BrainAnonymous y9EilyNessuna valutazione finora
- 4 Myonuclei Acquired by Overload Exercise Precede Hypertrophy and Are Not Lost On DetrainingDocumento6 pagine4 Myonuclei Acquired by Overload Exercise Precede Hypertrophy and Are Not Lost On DetrainingAline KreiNessuna valutazione finora
- A Promising Strategy To Promote RemyelinationDocumento7 pagineA Promising Strategy To Promote RemyelinationYNessuna valutazione finora
- Fructus SchisandraeDocumento14 pagineFructus SchisandraeHenrique Santa Capita CerqueiraNessuna valutazione finora
- 2022 Article 1233Documento24 pagine2022 Article 1233orellanaroco.10Nessuna valutazione finora
- Bayer 2018Documento38 pagineBayer 2018Lauda SinghNessuna valutazione finora
- Joa 1943 0323Documento12 pagineJoa 1943 0323Bernardo SteinbergNessuna valutazione finora
- Accepted Manuscript: Biochemical PharmacologyDocumento53 pagineAccepted Manuscript: Biochemical PharmacologyStanford LeeNessuna valutazione finora
- Mechanism Induced TrainingDocumento15 pagineMechanism Induced Trainingmarina robertaNessuna valutazione finora
- Remodeling of The Skeletal Muscle and Postsynaptic Component After Short Term Joint Immobilization and Aquatic TrainingDocumento8 pagineRemodeling of The Skeletal Muscle and Postsynaptic Component After Short Term Joint Immobilization and Aquatic TrainingRicardo Pereira NevesNessuna valutazione finora
- 137Documento9 pagine137Conciencia CristalinaNessuna valutazione finora
- Muhammad Salman Bilal: Operations ManagementDocumento22 pagineMuhammad Salman Bilal: Operations ManagementShamyl Farooq KhanNessuna valutazione finora
- Agile Supply Chain Management On FMCGDocumento26 pagineAgile Supply Chain Management On FMCGShamyl Farooq KhanNessuna valutazione finora
- Agile Meets Assessments Case Study On How To Do AgDocumento17 pagineAgile Meets Assessments Case Study On How To Do AgShamyl Farooq KhanNessuna valutazione finora
- Types of ResearchDocumento22 pagineTypes of ResearchShamyl Farooq KhanNessuna valutazione finora
- Codex Standard For Pickled Cucumbers (Cucumber Pickles) CODEX STAN 115-1981Documento11 pagineCodex Standard For Pickled Cucumbers (Cucumber Pickles) CODEX STAN 115-1981Shamyl Farooq KhanNessuna valutazione finora
- Intermittent Fasting: The Choice For A Healthier LifestyleDocumento11 pagineIntermittent Fasting: The Choice For A Healthier LifestyleShamyl Farooq KhanNessuna valutazione finora
- Best Ice Cream Business Plan WTH Financials PDFDocumento11 pagineBest Ice Cream Business Plan WTH Financials PDFShamyl Farooq KhanNessuna valutazione finora
- Validation of Reference Genes For QPCR Analysis of Resistance Training and Androgenic Anabolic Steroids On Hypothalamus Adrenal GL 2157 7536 1000180Documento7 pagineValidation of Reference Genes For QPCR Analysis of Resistance Training and Androgenic Anabolic Steroids On Hypothalamus Adrenal GL 2157 7536 1000180Shamyl Farooq KhanNessuna valutazione finora
- Structure of A Phosphoglycerate Mutase3-PhosphoglyDocumento6 pagineStructure of A Phosphoglycerate Mutase3-PhosphoglyShamyl Farooq KhanNessuna valutazione finora
- 10 Ep 4 2012-11Documento15 pagine10 Ep 4 2012-11Shamyl Farooq KhanNessuna valutazione finora
- Validation of Reference Genes For QPCR Analysis of Resistance Training and Androgenic Anabolic Steroids On Hypothalamus Adrenal GL 2157 7536 1000180Documento7 pagineValidation of Reference Genes For QPCR Analysis of Resistance Training and Androgenic Anabolic Steroids On Hypothalamus Adrenal GL 2157 7536 1000180Shamyl Farooq KhanNessuna valutazione finora
- Muscle Memory New Cellular Model For Atrophy Hypertrophy 16 PDFDocumento8 pagineMuscle Memory New Cellular Model For Atrophy Hypertrophy 16 PDFPavel Tony Jauregui AlorNessuna valutazione finora
- Pepsi CODocumento17 paginePepsi COShamyl Farooq KhanNessuna valutazione finora
- PLSC 102 - Course Outline - February2018Documento2 paginePLSC 102 - Course Outline - February2018Shamyl Farooq KhanNessuna valutazione finora
- Nuclie ArticleDocumento7 pagineNuclie ArticleShamyl Farooq KhanNessuna valutazione finora
- UNDP-PK-DGU-Federalism 18 Amendment Report-2012 PDFDocumento58 pagineUNDP-PK-DGU-Federalism 18 Amendment Report-2012 PDFShamyl Farooq Khan100% (1)
- Structure of A Phosphoglycerate Mutase3-PhosphoglyDocumento6 pagineStructure of A Phosphoglycerate Mutase3-PhosphoglyShamyl Farooq KhanNessuna valutazione finora
- Quranic Verses and Hadith Against Bribery and CorruptionDocumento3 pagineQuranic Verses and Hadith Against Bribery and CorruptionShamyl Farooq KhanNessuna valutazione finora
- Structure of A Phosphoglycerate Mutase3-PhosphoglyDocumento6 pagineStructure of A Phosphoglycerate Mutase3-PhosphoglyShamyl Farooq KhanNessuna valutazione finora
- 116 Wing Tsun Wooden Dummy TechniquesDocumento119 pagine116 Wing Tsun Wooden Dummy Techniquesdewdzz100% (54)
- The Risk of Hormone Anabolic Steroids and Oral Contraceptives Consumption in The Young 2329 6631.1000113Documento4 pagineThe Risk of Hormone Anabolic Steroids and Oral Contraceptives Consumption in The Young 2329 6631.1000113Shamyl Farooq KhanNessuna valutazione finora
- Advanced Home WorkoutDocumento1 paginaAdvanced Home WorkoutShamyl Farooq KhanNessuna valutazione finora
- Morphology of Prokaryotes & EukaryotesDocumento21 pagineMorphology of Prokaryotes & EukaryotesDhanush VNessuna valutazione finora
- The Epidermis in Wound HealingDocumento404 pagineThe Epidermis in Wound HealingongchenguanNessuna valutazione finora
- Stages of Growth and Development: Before BirthDocumento3 pagineStages of Growth and Development: Before BirthZowi Sian MedesNessuna valutazione finora
- NE LINKAGE - Test QuestionsDocumento9 pagineNE LINKAGE - Test QuestionsJacobmmcdonald0% (1)
- Dwnload Full Campbell Biology 10th Edition Reece Solutions Manual PDFDocumento36 pagineDwnload Full Campbell Biology 10th Edition Reece Solutions Manual PDFclintonabalosfxz100% (15)
- Total RBC Count by Hemocytometer - Hematology PracticalsDocumento10 pagineTotal RBC Count by Hemocytometer - Hematology Practicalsrahayu_jatiningsihNessuna valutazione finora
- Pharmacology: A Career inDocumento2 paginePharmacology: A Career insnikt7863443Nessuna valutazione finora
- Haploid ProductionDocumento25 pagineHaploid Productionitube100% (2)
- Mitosis Internet LessonDocumento3 pagineMitosis Internet LessonGLENMAR VINEGASNessuna valutazione finora
- Bohnert 2013Documento42 pagineBohnert 2013marianaNessuna valutazione finora
- Hormones As Signals, Classification of HormonesDocumento22 pagineHormones As Signals, Classification of HormonesHarini BalasubramanianNessuna valutazione finora
- Avian Influenza in ChickensDocumento19 pagineAvian Influenza in ChickensShah NawazNessuna valutazione finora
- Principle of DNA MicroarrayDocumento5 paginePrinciple of DNA MicroarrayDipteemaya Biswal100% (1)
- Simple R Tools For Genetic Markers ResearchDocumento3 pagineSimple R Tools For Genetic Markers ResearchIJAR JOURNALNessuna valutazione finora
- Plasmodium VivaxDocumento7 paginePlasmodium VivaxrobiahanitasariNessuna valutazione finora
- NBRI MarigoldDocumento2 pagineNBRI MarigoldShailendra RajanNessuna valutazione finora
- Science 4: TH NDDocumento3 pagineScience 4: TH NDNashaNessuna valutazione finora
- Reaction Paper On The Real EveDocumento2 pagineReaction Paper On The Real EveJoy Marisol Flores ManiaulNessuna valutazione finora
- Environment EcologyDocumento13 pagineEnvironment EcologyKa IfiNessuna valutazione finora
- 1960 Mower LearningtheoryDocumento585 pagine1960 Mower Learningtheoryfatty_mvNessuna valutazione finora
- IUCN Protected Areas Categories SystemDocumento2 pagineIUCN Protected Areas Categories SystemBharath SandeepNessuna valutazione finora
- BiodiversityDocumento25 pagineBiodiversityOrane CassanovaNessuna valutazione finora
- Organic Molecules Assignment MenuDocumento3 pagineOrganic Molecules Assignment Menuapi-434671855Nessuna valutazione finora
- Personal Data:: Curiculum VitaeDocumento7 paginePersonal Data:: Curiculum Vitaebudi darmantaNessuna valutazione finora
- Reproductive Medicine and Assisted Reproductive Technologies (A.R.T.)Documento23 pagineReproductive Medicine and Assisted Reproductive Technologies (A.R.T.)Azhari AhmadNessuna valutazione finora
- GM Food (IB MYP Essay)Documento10 pagineGM Food (IB MYP Essay)Trippin OnAcidNessuna valutazione finora
- HEMA Lec 02 Hematopoiesis 2Documento5 pagineHEMA Lec 02 Hematopoiesis 2Patrick AbellaNessuna valutazione finora
- ICSE Class 10 Biology 2007Documento7 pagineICSE Class 10 Biology 2007SACHIDANANDA SNessuna valutazione finora
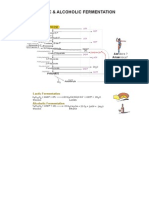
- Lactic and Alcoholic FermentationDocumento1 paginaLactic and Alcoholic FermentationDr. SHIVA AITHALNessuna valutazione finora
- Chen2014 PDFDocumento37 pagineChen2014 PDFJL Plantae ZCNessuna valutazione finora