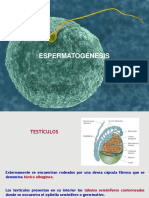
Potrebbero piacerti anche
- Clase 2 ObstetriciaDocumento47 pagineClase 2 ObstetriciaRaúl Alexis Menudo ProblemaNessuna valutazione finora
- Trabajo IvanDocumento12 pagineTrabajo IvanNatalia SáenzNessuna valutazione finora
- Cuestionario 8Documento3 pagineCuestionario 8Victor AngelNessuna valutazione finora
- Sesión 03 - Citología ExfoliativaDocumento10 pagineSesión 03 - Citología Exfoliativafreddy.quisperuedaNessuna valutazione finora
- Aparato Reproductor FemeninoDocumento17 pagineAparato Reproductor FemeninoJuan Esteban GarciaNessuna valutazione finora
- E1 - Ciclo Ovarico y Endometrial 2017Documento10 pagineE1 - Ciclo Ovarico y Endometrial 2017Marcelino GaravitoNessuna valutazione finora
- Cuadernillo Histologia B COMPLETODocumento459 pagineCuadernillo Histologia B COMPLETOALdiiCge100% (2)
- Ovogénesis 1Documento7 pagineOvogénesis 1catalinaNessuna valutazione finora
- Fisiologia de La Reproducción (Zootecnia)Documento24 pagineFisiologia de La Reproducción (Zootecnia)Jhon brandon Álvarez SolísNessuna valutazione finora
- Aparato Reproductor HembraDocumento46 pagineAparato Reproductor HembraGonzalo MunNessuna valutazione finora
- Ovo GenesisDocumento32 pagineOvo Genesisalejandra sanchezNessuna valutazione finora
- Ciclo MenstrualDocumento48 pagineCiclo Menstrualjaneth nacarino sanchezNessuna valutazione finora
- Guia EmbrioDocumento150 pagineGuia EmbrioLarissa ManziNessuna valutazione finora
- Fecundacinsegmentacineimplantacin 121206152556 Phpapp02Documento55 pagineFecundacinsegmentacineimplantacin 121206152556 Phpapp02María Esperanza Carrizo GarcíaNessuna valutazione finora
- Aparato Reproductor FemeninoDocumento20 pagineAparato Reproductor FemeninoKoa NunchiNessuna valutazione finora
- Modulo C IndiraDocumento5 pagineModulo C IndiraAngel BatistaNessuna valutazione finora
- Atividade Ciclo OvaricoDocumento4 pagineAtividade Ciclo OvaricoEster EmanuelyNessuna valutazione finora
- OVOGENESISDocumento10 pagineOVOGENESISLuis Angel Garcia ColinesNessuna valutazione finora
- Maduracion Del Ovocito, Fecundación y Desarrollo Embrionario Equipo 2Documento65 pagineMaduracion Del Ovocito, Fecundación y Desarrollo Embrionario Equipo 2Tania GuerraNessuna valutazione finora
- Fecundacion AlteraciónesDocumento57 pagineFecundacion AlteraciónesMelisaNessuna valutazione finora
- Libro Produccion de Huevos Cap1 El Huevo de GallinaDocumento28 pagineLibro Produccion de Huevos Cap1 El Huevo de Gallinafernie3007Nessuna valutazione finora
- Ciclo Ovárico HistologiaDocumento3 pagineCiclo Ovárico HistologiaMarier BermudezNessuna valutazione finora
- Ovogénesis y EspermatogénesisDocumento29 pagineOvogénesis y EspermatogénesisKevin BetancurNessuna valutazione finora
- Resumen LangmanDocumento20 pagineResumen LangmanGabrielaRojas60% (5)
- Tema 2 - Ciclo Ovarico Fisiologia de La Ovulacion y La MenstruacionDocumento11 pagineTema 2 - Ciclo Ovarico Fisiologia de La Ovulacion y La Menstruacioncarmen pastranaNessuna valutazione finora
- Histología Del Aparato Genital FemeninoDocumento8 pagineHistología Del Aparato Genital FemeninoIsbelia Valentina Rivas MolinaNessuna valutazione finora
- Ciclo Reproductor FemeninoDocumento9 pagineCiclo Reproductor FemeninoTefy Tullume100% (1)
- Ciclo Menstrual PPT FinalDocumento17 pagineCiclo Menstrual PPT FinalKevin Angelo Arteaga QuispeNessuna valutazione finora
- Desarrollo Folicular y OvulaciónDocumento16 pagineDesarrollo Folicular y OvulaciónHeinter E Villanueva-VelardeNessuna valutazione finora
- Reporte de Laboratorio de GametogénesisDocumento14 pagineReporte de Laboratorio de GametogénesisYessy Ballesteros100% (1)
- Lección No 1. Fisiología de La Reproduccion1Documento29 pagineLección No 1. Fisiología de La Reproduccion1vimarjuNessuna valutazione finora
- Paso 2 Norida OrdoñezDocumento16 paginePaso 2 Norida OrdoñezLiliana OrdoñezNessuna valutazione finora
- Ud 1 Parte 2Documento44 pagineUd 1 Parte 2Ana LopezNessuna valutazione finora
- Embriologia BasicaDocumento22 pagineEmbriologia BasicaJose AvilaNessuna valutazione finora
- Fisiología Femenina Antes Del Embarazo y Hormonas FemeninasDocumento10 pagineFisiología Femenina Antes Del Embarazo y Hormonas Femeninasnicolas100% (2)
- Monografia Desarrollo PrenatalDocumento42 pagineMonografia Desarrollo PrenatalICASTICA33% (3)
- Embrioapuntes (Parte I) - Diego MeléndezDocumento16 pagineEmbrioapuntes (Parte I) - Diego Meléndezdiego melendezNessuna valutazione finora
- Ciclo SexualDocumento31 pagineCiclo SexualSarai Vega HoyosNessuna valutazione finora
- Ensayo Ovarios y FoliculogènesisDocumento3 pagineEnsayo Ovarios y FoliculogènesisLiLi AltamiranoNessuna valutazione finora
- EspermatogénesisDocumento53 pagineEspermatogénesisAmanda Hernández CuadraNessuna valutazione finora
- Resumen Langman Apuntes CompletoDocumento21 pagineResumen Langman Apuntes CompletoL J0% (1)
- El Pibe Resumen de FecundaciónDocumento29 pagineEl Pibe Resumen de FecundaciónAntonella Violo100% (1)
- Fabricio EmbrioDocumento9 pagineFabricio EmbrioFabri IbañezNessuna valutazione finora
- Aparato Reproductor FemeninoDocumento11 pagineAparato Reproductor FemeninoMIREYA GARCIA NIETONessuna valutazione finora
- Ciclos Reproductores y Continuos PDFDocumento3 pagineCiclos Reproductores y Continuos PDFKlaudiia VenegasNessuna valutazione finora
- Implantación Embriogenesis y Desarrollo PlacentarioDocumento47 pagineImplantación Embriogenesis y Desarrollo PlacentariojeffelwarNessuna valutazione finora
- Meta1.3:VILLA LOPEZDocumento8 pagineMeta1.3:VILLA LOPEZFlor Denisse Villa LópezNessuna valutazione finora
- Ovogenesis y FoliculogenesisDocumento41 pagineOvogenesis y FoliculogenesisArturo Athie Athie100% (1)
- Fisiologia Del Ciclo MenstrualDocumento4 pagineFisiologia Del Ciclo MenstrualCristina RuizNessuna valutazione finora
- Primera Semana de Desarrollo EmbrionarioDocumento5 paginePrimera Semana de Desarrollo EmbrionarioEnoc GonzálezNessuna valutazione finora
- Loe11 FisiologiaDocumento30 pagineLoe11 FisiologiaJuan Pablo Giraldo MontoyaNessuna valutazione finora
- Fisiología de La GestaciónDocumento10 pagineFisiología de La Gestaciónrociofigueroa.m130Nessuna valutazione finora
- Etapa Concepcional CDDocumento10 pagineEtapa Concepcional CDKatherine TorresNessuna valutazione finora
- Embriologia General.Documento9 pagineEmbriologia General.Candela Carrion BustosNessuna valutazione finora
- 9 Resumen - Síntesis - Unidad 3 - Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital FemeninoDocumento17 pagine9 Resumen - Síntesis - Unidad 3 - Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital FemeninoMiranda LeonettiNessuna valutazione finora
- Embrio M1.4Documento15 pagineEmbrio M1.4Hector Efrain Garcia ChavezNessuna valutazione finora
- Fisiologia Del Eje H-H-ODocumento18 pagineFisiologia Del Eje H-H-OLuz Milagritos100% (1)
- Capitulo 23 Sistema Genital FemeninoDocumento16 pagineCapitulo 23 Sistema Genital FemeninoJohan ChoqueNessuna valutazione finora
- Ciclo Ovarico y EndometrialDocumento28 pagineCiclo Ovarico y EndometrialJuan Carlos Caldera MurilloNessuna valutazione finora
- MF0712_1 - Operaciones auxiliares en reproducción ganaderaDa EverandMF0712_1 - Operaciones auxiliares en reproducción ganaderaNessuna valutazione finora
- Giráldez, F. J. (Et Al.) 2003 Alimentación Del Ovino de Leche...Documento6 pagineGiráldez, F. J. (Et Al.) 2003 Alimentación Del Ovino de Leche...Ronald MQNessuna valutazione finora
- Vigilancia EpidemiologicaDocumento29 pagineVigilancia EpidemiologicaRonald MQNessuna valutazione finora
- 2019 Nestor Julian CardonaDocumento49 pagine2019 Nestor Julian CardonaRonald MQNessuna valutazione finora
- Fisiologia Animal MetabolismoDocumento16 pagineFisiologia Animal MetabolismoMónicaPBNessuna valutazione finora
- Ejercicos ADDocumento79 pagineEjercicos ADestudiosuja74Nessuna valutazione finora
- Estadística Ii: Lic. Jessica Elizabeth Chalco Suárez Lic. Wilbert Colque CandiaDocumento28 pagineEstadística Ii: Lic. Jessica Elizabeth Chalco Suárez Lic. Wilbert Colque CandiaDanitza D. OmarNessuna valutazione finora
- Proyecto Final de Laboratorio BILLARDocumento10 pagineProyecto Final de Laboratorio BILLARLuis Andres Ariza Osorio100% (1)
- Arboles Rojos Y NegrosDocumento8 pagineArboles Rojos Y Negrosjcastiblanco84Nessuna valutazione finora
- Solucionario de Tarea de Derivadas de Orden SuperiorDocumento5 pagineSolucionario de Tarea de Derivadas de Orden SuperiorJottaa YsNessuna valutazione finora
- Memoria Descriptiva. AquilinaDocumento2 pagineMemoria Descriptiva. AquilinaRaul Espinoza VargasNessuna valutazione finora
- Taller N 1 - Estabilidad en Los Sistemas Electricos de PotenciaDocumento6 pagineTaller N 1 - Estabilidad en Los Sistemas Electricos de PotenciaCarolCristinaGallegoLozanoNessuna valutazione finora
- Manual Osiloscopio OWONDocumento111 pagineManual Osiloscopio OWONJorge Luis Espin Claudio100% (3)
- Práctica Calificada N°2 - 2023-02 - 295716 - ESTADÍSTICA GENERAL - 2023-02 - FC-VIRCOM02D01MDocumento11 paginePráctica Calificada N°2 - 2023-02 - 295716 - ESTADÍSTICA GENERAL - 2023-02 - FC-VIRCOM02D01MNicollth CTNessuna valutazione finora
- PancreatitisDocumento13 paginePancreatitisluis ivanNessuna valutazione finora
- Diseà o de Voladura de RocasDocumento79 pagineDiseà o de Voladura de RocasYenny MarcelaNessuna valutazione finora
- Trazo en PlantaDocumento22 pagineTrazo en PlantaAngelica Rosa100% (1)
- Taller # 1 Calculo IntegralDocumento1 paginaTaller # 1 Calculo IntegralhernanescorciaNessuna valutazione finora
- Pruebas Saber - Naturales 4Documento1 paginaPruebas Saber - Naturales 4victorandy2002Nessuna valutazione finora
- Lenguaje Musical 1°Documento8 pagineLenguaje Musical 1°Marco AntonioNessuna valutazione finora
- Module 9 Pillar GridingDocumento4 pagineModule 9 Pillar GridingEdgar A Villegas ANessuna valutazione finora
- PCM, DPCM, DM, AdmDocumento2 paginePCM, DPCM, DM, AdmDavidNessuna valutazione finora
- Clase CubicacionesDocumento32 pagineClase CubicacionesNicolás Matías Castillo Morales100% (1)
- Calcbosk-User-Manual Final 12202018 EspDocumento69 pagineCalcbosk-User-Manual Final 12202018 EspSergio MoralesNessuna valutazione finora
- Evaluacion Avanzada 6Documento3 pagineEvaluacion Avanzada 6Ampa CordubaNessuna valutazione finora
- UNIDAD 1 PASO 1 Planeacion Estrategica 2019Documento5 pagineUNIDAD 1 PASO 1 Planeacion Estrategica 2019Andrea MuñozNessuna valutazione finora
- Elementos AntiserruchosDocumento3 pagineElementos AntiserruchosPaul Kevin Bautista Palacios100% (2)
- Teorema de BayesDocumento18 pagineTeorema de BayesMaria MachadoNessuna valutazione finora
- Problemas de Numeros Primos y Compuestos para Cuarto de SecundariaDocumento3 pagineProblemas de Numeros Primos y Compuestos para Cuarto de SecundariaTito VillafaneNessuna valutazione finora
- Práctica 2Documento4 paginePráctica 2Eliecer CardenasNessuna valutazione finora
- Tercer Examen de FísicaDocumento9 pagineTercer Examen de FísicaDayana ArqueNessuna valutazione finora
- Actividad 5 FisicaDocumento5 pagineActividad 5 FisicaJuan Rulfo75% (4)
- Datacion RadiometricaDocumento4 pagineDatacion RadiometricaDiana Liz Lozano FloresNessuna valutazione finora
- Grado 6. Guia 5-GeometriaDocumento7 pagineGrado 6. Guia 5-GeometriaHernando Cuestatic50% (2)
- Actividad Caida Libre Realizada.Documento3 pagineActividad Caida Libre Realizada.Dorina CardenasNessuna valutazione finora