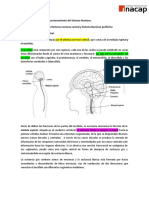
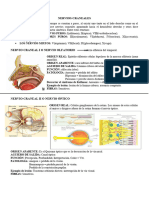
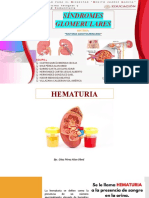
Potrebbero piacerti anche
- El cannabis en patologías del sistema nervioso centralDa EverandEl cannabis en patologías del sistema nervioso centralNessuna valutazione finora
- Anatomía y Fisiología del Sistema NerviosoDa EverandAnatomía y Fisiología del Sistema NerviosoValutazione: 4 su 5 stelle4/5 (19)
- Anatomia y Fisiología del Sistema Nervioso IIDa EverandAnatomia y Fisiología del Sistema Nervioso IIValutazione: 4 su 5 stelle4/5 (28)
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Da EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Nessuna valutazione finora
- 6 Sistema NerviosoDocumento54 pagine6 Sistema NerviosoXavi Moran100% (1)
- Documento Modulo I Parte 2Documento8 pagineDocumento Modulo I Parte 2Victoria Lucia LopezNessuna valutazione finora
- Sistema Nervioso Cuestionario 1Documento14 pagineSistema Nervioso Cuestionario 1Maria Gabriela MendozaNessuna valutazione finora
- Bases Biologicas de La Conducta.Documento12 pagineBases Biologicas de La Conducta.Krisbely RangelNessuna valutazione finora
- Sistema NerviosoDocumento73 pagineSistema NerviosoVeronica Pineda100% (1)
- Sinopsis Del CerebroDocumento3 pagineSinopsis Del CerebroDaniel Aybar CarreraNessuna valutazione finora
- Sistema Nervioso Indice 1Documento23 pagineSistema Nervioso Indice 1Erick Morales GonzalezNessuna valutazione finora
- Sistema Nervioso - Anatomía Autor Avelina Tortosa MorenoDocumento31 pagineSistema Nervioso - Anatomía Autor Avelina Tortosa MorenoVerónica Cruz MoralesNessuna valutazione finora
- (Haines) Introducción A La Estructura Del Sistema Nervioso CentralDocumento5 pagine(Haines) Introducción A La Estructura Del Sistema Nervioso CentralCesar EgusquizaNessuna valutazione finora
- U6 Bio Resumen Re Mil Terminado IiiiiiujuDocumento21 pagineU6 Bio Resumen Re Mil Terminado Iiiiiiujubianca dileoNessuna valutazione finora
- Neuroanatomía-Curso Online PUCDocumento120 pagineNeuroanatomía-Curso Online PUCLauraguilar100% (8)
- Guia 4 ListaDocumento13 pagineGuia 4 ListaYazmin loraNessuna valutazione finora
- Atencion de Enfermeria A Pacientes Con Alteraciones NeurologicasDocumento30 pagineAtencion de Enfermeria A Pacientes Con Alteraciones NeurologicasAgny MoraNessuna valutazione finora
- Guía Teorica y Practica de Fisiopalogía 2020 (1) - Páginas-172-282Documento111 pagineGuía Teorica y Practica de Fisiopalogía 2020 (1) - Páginas-172-282Nahx JkNessuna valutazione finora
- Neuroanatomía. Dr. Andrés González GarridoDocumento26 pagineNeuroanatomía. Dr. Andrés González Garridojose jaime botello valle100% (1)
- Anatomia PsicologiaDocumento6 pagineAnatomia PsicologiaKaren Allendes Raffernau100% (1)
- Resumen Neurobiología FINALDocumento98 pagineResumen Neurobiología FINALCynthia ScottiNessuna valutazione finora
- Tarea 6 y 7. Anat. Fisiol Del Sistema Nervioso.Documento7 pagineTarea 6 y 7. Anat. Fisiol Del Sistema Nervioso.Janice Fígaro SimonsNessuna valutazione finora
- Resumen Final Neuro FerreDocumento22 pagineResumen Final Neuro FerreMili NunesNessuna valutazione finora
- Organización Estructural y Funcional Del SNCDocumento14 pagineOrganización Estructural y Funcional Del SNCjesus davidNessuna valutazione finora
- Sistema Nervioso IDocumento16 pagineSistema Nervioso Ilinda flor camarena muñozNessuna valutazione finora
- Sistema NerviosoDocumento10 pagineSistema NerviosoNicoNessuna valutazione finora
- ANATOMOFISILOGÍA II - Resumen para El FinalDocumento85 pagineANATOMOFISILOGÍA II - Resumen para El Finalcandela rodriguez gherbazNessuna valutazione finora
- Sistema Nervioso SimpaticoDocumento20 pagineSistema Nervioso SimpaticoLina VelezNessuna valutazione finora
- Crecimiento y Desarrollo UP7Documento21 pagineCrecimiento y Desarrollo UP7Franco LibertoNessuna valutazione finora
- M1 T6 - NeuroPsDocumento13 pagineM1 T6 - NeuroPsThonny EspinosaNessuna valutazione finora
- Organización Funcional Del Sistema NerviosoDocumento6 pagineOrganización Funcional Del Sistema NerviosoMarcela Gómez ChacónNessuna valutazione finora
- Resumen Fina Parte 1Documento63 pagineResumen Fina Parte 1Maria Celina MassaNessuna valutazione finora
- Funciones Del Sistema Nervioso Central y PeriféricoDocumento6 pagineFunciones Del Sistema Nervioso Central y PerifériconadiaNessuna valutazione finora
- Investigacion (SISTEMA NERVIOSO)Documento5 pagineInvestigacion (SISTEMA NERVIOSO)LuiGabo DunsNessuna valutazione finora
- Las Neurociencias 2Documento33 pagineLas Neurociencias 2Marco DíazNessuna valutazione finora
- Apuntes NeuroanatomíaDocumento109 pagineApuntes NeuroanatomíaMichelle Acuña EspinozaNessuna valutazione finora
- Guia 1 GenerneuroanatoDocumento28 pagineGuia 1 GenerneuroanatoBerenice TorresNessuna valutazione finora
- Plan de Prevención y Control de Patologías de Origen LaboralDocumento19 paginePlan de Prevención y Control de Patologías de Origen LaboralAlexander Mejia Navarro100% (1)
- Ev 4.1Documento12 pagineEv 4.1Hanny DavilaNessuna valutazione finora
- Sistema NerviosoDocumento2 pagineSistema NerviosoK Susana M Bazan100% (1)
- NEUROLOGIADocumento10 pagineNEUROLOGIAMarcos Manuel Vásquez MendozaNessuna valutazione finora
- Zuleica 1 AntmiaDocumento6 pagineZuleica 1 AntmiaDiover PaulinoNessuna valutazione finora
- Trabajo de PsicobiologiaDocumento11 pagineTrabajo de PsicobiologiaJ Mora GañanNessuna valutazione finora
- Sistema Nervioso. GeneralidadesDocumento59 pagineSistema Nervioso. Generalidadesgwbn6qhgt9Nessuna valutazione finora
- NeuroanatomiaDocumento30 pagineNeuroanatomiaDiego Arturo Muñoz SepúlvedaNessuna valutazione finora
- Atlas Del EncéfaloDocumento6 pagineAtlas Del EncéfaloLorena LeylerNessuna valutazione finora
- 07-15-2023 203059 PM Características de Los Sistemas Nerviosos Central y Periférico.Documento9 pagine07-15-2023 203059 PM Características de Los Sistemas Nerviosos Central y Periférico.Lourdes Cruz CastroNessuna valutazione finora
- Componentes Del Sistema NerviosoDocumento6 pagineComponentes Del Sistema NerviosoJazmin Martinez zambranoNessuna valutazione finora
- Presentación 3 Procesos de AprendizajeDocumento38 paginePresentación 3 Procesos de AprendizajeJavier ValdezNessuna valutazione finora
- Sistema NerviosoDocumento8 pagineSistema NerviosoJuliana AcevedoNessuna valutazione finora
- Resumen GeneralDocumento19 pagineResumen Generalabigail tonelottoNessuna valutazione finora
- Diviana Rocca Seccion B Neurofisiologia y Fisiopatologia Unidad 3Documento114 pagineDiviana Rocca Seccion B Neurofisiologia y Fisiopatologia Unidad 3oriana roccaNessuna valutazione finora
- Alteraciones Del Sistema Nervioso Central PresentaciónDocumento31 pagineAlteraciones Del Sistema Nervioso Central PresentaciónLuis MartinezNessuna valutazione finora
- GeneralidadessistnervDocumento22 pagineGeneralidadessistnervJuan OjedaNessuna valutazione finora
- Resumen IIDocumento8 pagineResumen IICarla MálagaNessuna valutazione finora
- 6.15 Sistema Nervioso Central CerebroDocumento8 pagine6.15 Sistema Nervioso Central Cerebrojose davidNessuna valutazione finora
- Final de Psicofisiologia (Cap Del 6 Al 17)Documento21 pagineFinal de Psicofisiologia (Cap Del 6 Al 17)Micaela HovenNessuna valutazione finora
- Resumen de Generalidades Del Sistema Nervioso.Documento7 pagineResumen de Generalidades Del Sistema Nervioso.Adriana ClarosNessuna valutazione finora
- Esquema Sistema Nervioso CentralDocumento7 pagineEsquema Sistema Nervioso CentralAngecrissNessuna valutazione finora
- Sistema Nervioso PerifericoDocumento4 pagineSistema Nervioso PerifericoCarlos Proaño SalmonNessuna valutazione finora
- Iiip - Clase 2 - Penicilinas Naturales y SemisinteticasDocumento43 pagineIiip - Clase 2 - Penicilinas Naturales y SemisinteticasPedro NavarroNessuna valutazione finora
- IIIP - Clase 3 - CefalosporinasDocumento26 pagineIIIP - Clase 3 - CefalosporinasPedro NavarroNessuna valutazione finora
- 03 Reconocimiento Unidad 1 - Farmacologia Del Sistema NerviosoDocumento12 pagine03 Reconocimiento Unidad 1 - Farmacologia Del Sistema NerviosoLeandro Jose Marrugo OspinoNessuna valutazione finora
- Patologia Ano RectoDocumento14 paginePatologia Ano RectoPedro NavarroNessuna valutazione finora
- Patologia Ano RectoDocumento14 paginePatologia Ano RectoPedro NavarroNessuna valutazione finora
- Trastorno Del Crecimiento y Diferenciación CelularDocumento9 pagineTrastorno Del Crecimiento y Diferenciación CelularPedro NavarroNessuna valutazione finora
- Historia Natural de Tetralogia de FallotDocumento1 paginaHistoria Natural de Tetralogia de FallotEloisa Domínguez89% (9)
- TuberculosisDocumento7 pagineTuberculosisdark- shinigamiNessuna valutazione finora
- Infografía Conjuntivitis LeñosaDocumento1 paginaInfografía Conjuntivitis Leñosacarol garciaNessuna valutazione finora
- Manejo HidroelectroliticoDocumento59 pagineManejo HidroelectroliticoJheremi Samir Rafael MaurateNessuna valutazione finora
- Caso ClinicoDocumento26 pagineCaso ClinicoMark100% (1)
- Preguntas Cirugía GeneralDocumento10 paginePreguntas Cirugía GeneralLuis Álvaro Castillo CerveraNessuna valutazione finora
- Reacciones de HipersensibilidadDocumento2 pagineReacciones de HipersensibilidadKaren Lizeth Hernández JaimeNessuna valutazione finora
- 1477 7827 8 21.en - EsDocumento7 pagine1477 7827 8 21.en - EsJosé Manuel OsortoNessuna valutazione finora
- Caso Clínico 9. Cocos Gram Negativos - NeisseriaDocumento4 pagineCaso Clínico 9. Cocos Gram Negativos - NeisseriaGris GuevNessuna valutazione finora
- Cactus GrandiflorusDocumento4 pagineCactus GrandiflorusLola MartinezNessuna valutazione finora
- SIMULACION (En Oftalmología)Documento41 pagineSIMULACION (En Oftalmología)Manuel Ángel Marcos FernándezNessuna valutazione finora
- Don T Touch Lesions - OficialDocumento40 pagineDon T Touch Lesions - OficialHans M. Valdiviezo BonifazNessuna valutazione finora
- Anemia Post HemorragicaDocumento8 pagineAnemia Post HemorragicaAlberto Delgado100% (1)
- Ictericia NeonatalDocumento8 pagineIctericia NeonatalErick Ortiz AvilésNessuna valutazione finora
- SX Hepatoesplenico. Pediatría II.Documento16 pagineSX Hepatoesplenico. Pediatría II.Alejandro TovarNessuna valutazione finora
- CASOS y Ejercicios HEPATOPATIAS 2022Documento9 pagineCASOS y Ejercicios HEPATOPATIAS 2022copiesteriaraquelNessuna valutazione finora
- Anfotericinas - Sociedad Argentina de InfectologiaDocumento14 pagineAnfotericinas - Sociedad Argentina de InfectologiaByron Miguel Cacay TorresNessuna valutazione finora
- Custionario PDFDocumento5 pagineCustionario PDFolga janeth cuellar urreaNessuna valutazione finora
- Clase 1 PropedéuticaDocumento33 pagineClase 1 PropedéuticaAnggy Maryury Molina MoncadaNessuna valutazione finora
- Trastornos Gonadales FisiopatologiaDocumento91 pagineTrastornos Gonadales Fisiopatologiasoot spritesNessuna valutazione finora
- Hematologia CompletoDocumento472 pagineHematologia CompletoNoelia Ballestas100% (4)
- M4-CSS X MapaDocumento1 paginaM4-CSS X MapaFederico MininNessuna valutazione finora
- Pares Craneales CompletoDocumento8 paginePares Craneales CompletoYaritza PerezNessuna valutazione finora
- USG Renal y PancreasDocumento125 pagineUSG Renal y PancreasGabriel HernándezNessuna valutazione finora
- Bronquiectasias SeparDocumento142 pagineBronquiectasias SeparJesus Mendez RodriguezNessuna valutazione finora
- Bacterias de Infecciones de Tejidos Blandos, Sistémicas y ZoonóticasDocumento10 pagineBacterias de Infecciones de Tejidos Blandos, Sistémicas y ZoonóticasMAIRA VANESA SANCHEZ RAMIREZNessuna valutazione finora
- Recurso 1Documento10 pagineRecurso 1Meenthal HealthNessuna valutazione finora
- Síndromes GlomerularesDocumento71 pagineSíndromes Glomerularesbrenda jazmin muñoz ramirezNessuna valutazione finora
- Introducción A La GastroenterologíaDocumento85 pagineIntroducción A La GastroenterologíaJose Peña Rivas100% (2)
- Clasificacion de Insuficiencia CardiacaDocumento8 pagineClasificacion de Insuficiencia CardiacaAlicia Ramírez HernándezNessuna valutazione finora