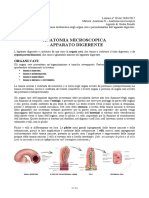
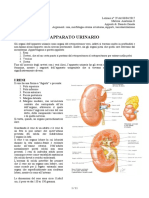
Potrebbero piacerti anche
- 16 - Anatomia II - 08.03.2017 - RDocumento9 pagine16 - Anatomia II - 08.03.2017 - RSandroNessuna valutazione finora
- 32 - Anatomia II - 3.05.2017Documento8 pagine32 - Anatomia II - 3.05.2017SandroNessuna valutazione finora
- 19 - Anatomia II - 15.03.2017 - RDocumento12 pagine19 - Anatomia II - 15.03.2017 - RSandroNessuna valutazione finora
- 41-Anatomia II - 25-05-2017 RDocumento9 pagine41-Anatomia II - 25-05-2017 RSandroNessuna valutazione finora
- 31 - Anatomia II - 27.04.2017 RDocumento10 pagine31 - Anatomia II - 27.04.2017 RSandroNessuna valutazione finora
- 05 - Anatomia II - 11.01.2017 - RDocumento6 pagine05 - Anatomia II - 11.01.2017 - RSandroNessuna valutazione finora
- 06 - Anatomia II - 16.01.2017Documento7 pagine06 - Anatomia II - 16.01.2017SandroNessuna valutazione finora
- 08 - Anatomia II - 18.01.2017 - RDocumento9 pagine08 - Anatomia II - 18.01.2017 - RSandroNessuna valutazione finora
- 05 - Anatomia II - 11.01.2017 - RDocumento6 pagine05 - Anatomia II - 11.01.2017 - RSandroNessuna valutazione finora
- 04 - Anatomia II - 09.01.2017Documento10 pagine04 - Anatomia II - 09.01.2017SandroNessuna valutazione finora
- 02 - Anatomia II - 05.12.2016 - RDocumento11 pagine02 - Anatomia II - 05.12.2016 - RSandroNessuna valutazione finora
- 30 - Anatomia II - 26-04-2017 - RDocumento11 pagine30 - Anatomia II - 26-04-2017 - RSandroNessuna valutazione finora
- 04 - Anatomia II - 09.01.2017Documento10 pagine04 - Anatomia II - 09.01.2017SandroNessuna valutazione finora
- 01 - Anatomia II - R 30.11.2016Documento8 pagine01 - Anatomia II - R 30.11.2016SandroNessuna valutazione finora
- 03 - Anatomia II - 07.12.2016Documento10 pagine03 - Anatomia II - 07.12.2016SandroNessuna valutazione finora
- 02 - Anatomia II - 05.12.2016 - RDocumento11 pagine02 - Anatomia II - 05.12.2016 - RSandroNessuna valutazione finora
- 27 - Anatomia II - 03.04.2017 RDocumento11 pagine27 - Anatomia II - 03.04.2017 RSandroNessuna valutazione finora
- 01 - Anatomia II - R 30.11.2016Documento8 pagine01 - Anatomia II - R 30.11.2016SandroNessuna valutazione finora
- 09 - Anatomia II - 20.02.2017Documento8 pagine09 - Anatomia II - 20.02.2017SandroNessuna valutazione finora
- 25 - Anatomia II - 29-03-2017 RDocumento7 pagine25 - Anatomia II - 29-03-2017 RSandroNessuna valutazione finora
- 28 - Anatomia II 05-04-2017 RDocumento11 pagine28 - Anatomia II 05-04-2017 RSandroNessuna valutazione finora
- 29 - Anatomia II - 06-04-2017 - RDocumento11 pagine29 - Anatomia II - 06-04-2017 - RSandroNessuna valutazione finora
- 28 - Anatomia II 05-04-2017 RDocumento11 pagine28 - Anatomia II 05-04-2017 RSandroNessuna valutazione finora
- 10 - Anatomia II - 22.02.2017 - RDocumento9 pagine10 - Anatomia II - 22.02.2017 - RSandroNessuna valutazione finora
- 12 - Anatomia II - 27.02.2017Documento12 pagine12 - Anatomia II - 27.02.2017SandroNessuna valutazione finora
- 11 - Anatomia II - 23.02.2017Documento11 pagine11 - Anatomia II - 23.02.2017SandroNessuna valutazione finora
- 13 - Anatomia II - 01.03.2017 - RDocumento14 pagine13 - Anatomia II - 01.03.2017 - RSandroNessuna valutazione finora
- 15 - Anatomia II - 06.03.2017Documento10 pagine15 - Anatomia II - 06.03.2017SandroNessuna valutazione finora
- 14 AnatomiaII 02.03.2017 RDocumento14 pagine14 AnatomiaII 02.03.2017 RSandroNessuna valutazione finora
- 16 - Anatomia II - 08.03.2017 - RDocumento9 pagine16 - Anatomia II - 08.03.2017 - RSandroNessuna valutazione finora
- 1.sistemi Sensoriali 01Documento23 pagine1.sistemi Sensoriali 01larisutzafNessuna valutazione finora
- Da Cosa Nasce Cosa1Documento15 pagineDa Cosa Nasce Cosa1Giulia PresenzaNessuna valutazione finora
- Anche Gli Animali Hanno Un'anima!Documento42 pagineAnche Gli Animali Hanno Un'anima!Daniele GullàNessuna valutazione finora
- Il Gusto Sapore, Profumo e AromaDocumento5 pagineIl Gusto Sapore, Profumo e AromaRiccardo MuccardoNessuna valutazione finora
- Sds Bortoli2016Documento12 pagineSds Bortoli2016aldo123456789Nessuna valutazione finora
- Somestesia PDFDocumento28 pagineSomestesia PDFVincenzo BinantiNessuna valutazione finora
- La Percezione Multisensoriale - Capitolo 1Documento6 pagineLa Percezione Multisensoriale - Capitolo 1Jury PoklukarNessuna valutazione finora
- Dalla Testa Ai PiediDocumento164 pagineDalla Testa Ai PiediBruno CapitolinoNessuna valutazione finora
- Controllo Del MovimentoDocumento25 pagineControllo Del MovimentoElle DownloadNessuna valutazione finora
- Rieducazione Funzionale PDFDocumento63 pagineRieducazione Funzionale PDFpaolo68audisioNessuna valutazione finora
- L'Ambiente Come Fattore Di Apprendimento Nella Scuola Dell'InfanziaDocumento21 pagineL'Ambiente Come Fattore Di Apprendimento Nella Scuola Dell'InfanziaElisa Lo MonacoNessuna valutazione finora
- Opuscolo Dr. PantaleoDocumento40 pagineOpuscolo Dr. PantaleoCipriano D'AmbrosioNessuna valutazione finora
- Stimoli PDFDocumento60 pagineStimoli PDFOvidiu RaduNessuna valutazione finora
- I Sensi - Percorso Didattico Classe 5aDocumento25 pagineI Sensi - Percorso Didattico Classe 5aCurricolo Barberino100% (8)
- Allenamento IdeomotorioDocumento11 pagineAllenamento IdeomotorioFilippo Eduardo AranitiNessuna valutazione finora
- Epiteli SensorialiDocumento5 pagineEpiteli Sensorialiapi-3704348100% (1)
- 1 - Sensibilità SomestesicaDocumento23 pagine1 - Sensibilità Somestesicaapi-3704348100% (3)
- Effetti Mentali e Psicologici Del MassaggioDocumento18 pagineEffetti Mentali e Psicologici Del MassaggioohananlbNessuna valutazione finora
- Guardare Pensare Progettare by Riccardo FalcinelliDocumento326 pagineGuardare Pensare Progettare by Riccardo Falcinelliangiolina gaudio100% (1)
- Sensazione - Percezione - ConsapevolezzaDocumento78 pagineSensazione - Percezione - Consapevolezzayellowflower_2Nessuna valutazione finora
- Introduzione Alla Psicologia PDFDocumento24 pagineIntroduzione Alla Psicologia PDFFrancesco Zanca100% (5)
- Comunicazione Del CaneDocumento8 pagineComunicazione Del CaneGianlucaCiroAttanasioNessuna valutazione finora
- 42-4 Televisione StereoscopicaDocumento14 pagine42-4 Televisione StereoscopicaRenzo SattiNessuna valutazione finora