Potrebbero piacerti anche
- Células eucariontes: estrutura e funçãoDocumento5 pagineCélulas eucariontes: estrutura e funçãoLucas NevesNessuna valutazione finora
- Fechamento Problema 2Documento4 pagineFechamento Problema 2anna luisaNessuna valutazione finora
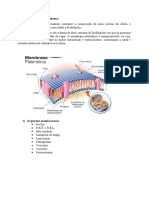
- Membrana CelularDocumento10 pagineMembrana CelularEsmeralda MuchisseNessuna valutazione finora
- Estrutura das Membranas CelularesDocumento19 pagineEstrutura das Membranas CelularesThiago SantosNessuna valutazione finora
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Documento13 pagineModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixNessuna valutazione finora
- Compartimentos Intracelulares e TransporteDocumento8 pagineCompartimentos Intracelulares e Transportefevabax924Nessuna valutazione finora
- Biologia - Membrana Celular e Transportes TransmembranaresDocumento5 pagineBiologia - Membrana Celular e Transportes TransmembranaresJoão AzevedoNessuna valutazione finora
- Biologia Molecular Da Célula (Alberts Et Al, 5 Ed.)Documento13 pagineBiologia Molecular Da Célula (Alberts Et Al, 5 Ed.)Rodrigo SimaoNessuna valutazione finora
- Resumo Biocel 1 Etapa - LuizaDocumento19 pagineResumo Biocel 1 Etapa - Luizaraf.marques.20Nessuna valutazione finora
- Estrutura e função das membranas celulares e sistemas de transporteDocumento14 pagineEstrutura e função das membranas celulares e sistemas de transporteLuis RicardoNessuna valutazione finora
- MI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaDocumento4 pagineMI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaEriczinNessuna valutazione finora
- Apostila-Citologia-pg 19 A 34Documento16 pagineApostila-Citologia-pg 19 A 34denisetg100% (2)
- BCELDocumento12 pagineBCELMaria lNessuna valutazione finora
- Membrana plasmática: estrutura e funçõesDocumento4 pagineMembrana plasmática: estrutura e funçõesDuda TavaresNessuna valutazione finora
- Resumo ProliferaçãoDocumento41 pagineResumo ProliferaçãoRodrigo Augusto100% (8)
- HISTOLOGIADocumento18 pagineHISTOLOGIARebecca SicariNessuna valutazione finora
- Aula MembranasDocumento33 pagineAula MembranasClaudio CirneNessuna valutazione finora
- Resumo - GenéticaDocumento16 pagineResumo - Genéticaisabeljulia968Nessuna valutazione finora
- Compartimentos Intracelulares e Endereçamento de ProteínasDocumento11 pagineCompartimentos Intracelulares e Endereçamento de ProteínasGabriel Da Silva SengesNessuna valutazione finora
- Membrana PlasmáticaDocumento10 pagineMembrana PlasmáticaNz NaganNessuna valutazione finora
- Membrana Plasmática: Transporte e EstruturasDocumento32 pagineMembrana Plasmática: Transporte e EstruturasPriscila MarinNessuna valutazione finora
- Histologia - CitologiaDocumento12 pagineHistologia - CitologiaPhelipe MonteiroNessuna valutazione finora
- Células: estrutura e função básicaDocumento33 pagineCélulas: estrutura e função básicaÂngela AlvesNessuna valutazione finora
- Fisiologia VegetalDocumento8 pagineFisiologia VegetalRafael GuardadoNessuna valutazione finora
- A estrutura e função da célulaDocumento8 pagineA estrutura e função da célulaKarina PezotiNessuna valutazione finora
- Estudo Biologia2Documento9 pagineEstudo Biologia2WQWQWEQWEQNessuna valutazione finora
- Membrana Celular: Transporte e Estrutura emDocumento46 pagineMembrana Celular: Transporte e Estrutura emTatiane ValeNessuna valutazione finora
- Obtenção de Matéria PDFDocumento15 pagineObtenção de Matéria PDFLeticia RussoNessuna valutazione finora
- Membrana celular: funções e estruturaDocumento20 pagineMembrana celular: funções e estruturaMario Maganizo JuniorNessuna valutazione finora
- Matéria Orgânica PDFDocumento13 pagineMatéria Orgânica PDFLeticia RussoNessuna valutazione finora
- Membrana Plasmática: Estrutura, Funções e TransporteDocumento5 pagineMembrana Plasmática: Estrutura, Funções e TransporteMoises FerroNessuna valutazione finora
- Biologia Celular pt1Documento22 pagineBiologia Celular pt1Rapha BuenoNessuna valutazione finora
- Resumo Complexo JuncionalDocumento5 pagineResumo Complexo JuncionalLuíza SiqueiraNessuna valutazione finora
- Apostila de Biologia - Vol 2Documento38 pagineApostila de Biologia - Vol 2Biologia Concurso Vestibular100% (2)
- Biologia Transporte Na CélulaDocumento3 pagineBiologia Transporte Na CélulaMárcio Simões GomesNessuna valutazione finora
- Citoesqueleto e CitoplasmaDocumento35 pagineCitoesqueleto e CitoplasmaRicardo GomesNessuna valutazione finora
- Princípios de Neurociência - Kandel 5a EdDocumento40 paginePrincípios de Neurociência - Kandel 5a EdcarlosNessuna valutazione finora
- CitoesqueletoDocumento4 pagineCitoesqueletoCláudia MartinsNessuna valutazione finora
- Biologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Documento65 pagineBiologia Celular e Genética - Aula 4 (Transporte, Organelas e Citoesqueleto)Leticia RodriguesNessuna valutazione finora
- Membrana Plasmática - BIOLOGIADocumento2 pagineMembrana Plasmática - BIOLOGIAJully ReisNessuna valutazione finora
- As células: unidades funcionais dos seres vivosDocumento10 pagineAs células: unidades funcionais dos seres vivosHálysson Nogueira ShalomNessuna valutazione finora
- Psicobiologia PsiclinicaDocumento86 paginePsicobiologia Psiclinicaneuropartilha4665Nessuna valutazione finora
- Biologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDocumento25 pagineBiologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDouglas NobreNessuna valutazione finora
- O papel dos filamentos do citoesqueletoDocumento8 pagineO papel dos filamentos do citoesqueletoAdamastor NetoNessuna valutazione finora
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDocumento7 pagineBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaNessuna valutazione finora
- Estudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasDocumento2 pagineEstudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasLuiza LopesNessuna valutazione finora
- Fichas - Organelas CitoplasmáticasDocumento6 pagineFichas - Organelas CitoplasmáticasEstefania RoblesNessuna valutazione finora
- Especializações Da Membrana PlasmáticaDocumento18 pagineEspecializações Da Membrana PlasmáticaPaula AlmeidaNessuna valutazione finora
- Resumos de Biologia CelularDocumento17 pagineResumos de Biologia CelularCaroline LimaNessuna valutazione finora
- Componentes celularesDocumento17 pagineComponentes celularesVera DominguesNessuna valutazione finora
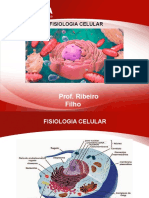
- Fisiologia celular: estrutura e função das célulasDocumento15 pagineFisiologia celular: estrutura e função das célulasAlícia Hellen100% (1)
- Membrana Celular: Características e FunçõesDocumento24 pagineMembrana Celular: Características e FunçõesAna Barth100% (1)
- A estrutura e função da membrana plasmáticaDocumento2 pagineA estrutura e função da membrana plasmáticaMaria Theresa MoreiraNessuna valutazione finora
- Membrana Plasmática. Digestão IntracelularDocumento27 pagineMembrana Plasmática. Digestão IntracelularBruno PereiraNessuna valutazione finora
- BiocelDocumento23 pagineBiocelSalvar Meus JoguinhosNessuna valutazione finora
- Gripes No MundoDocumento1 paginaGripes No MundoThais GonzagaNessuna valutazione finora
- Doenças Biologia1Documento2 pagineDoenças Biologia1ti18Nessuna valutazione finora
- Conselhos Úteis para Quem Vai Fazer A Cidadania Italiana Na Itália - Antes de Viajar Cidadania ItalianaDocumento3 pagineConselhos Úteis para Quem Vai Fazer A Cidadania Italiana Na Itália - Antes de Viajar Cidadania ItalianaThais GonzagaNessuna valutazione finora
- Trabalho Da Exposição Mario TestinoDocumento1 paginaTrabalho Da Exposição Mario TestinoThais GonzagaNessuna valutazione finora
- Cheesecake CopinhoDocumento1 paginaCheesecake CopinhoThais GonzagaNessuna valutazione finora
- Mousse de Chocolate fácilDocumento1 paginaMousse de Chocolate fácilThais GonzagaNessuna valutazione finora
- Atraente Alma-Brasileira FlautaDocumento4 pagineAtraente Alma-Brasileira FlautaThais GonzagaNessuna valutazione finora
- CandvagaDocumento7 pagineCandvagaAndreza Silveira PereiraNessuna valutazione finora
- SPM1202A caixa acústica ativa de alta potência e resposta planaDocumento1 paginaSPM1202A caixa acústica ativa de alta potência e resposta planaAlex DumontNessuna valutazione finora
- Medição de potência em circuitos elétricosDocumento22 pagineMedição de potência em circuitos elétricosVitor Fiorito da Silva0% (1)
- Uninter - Caderno de Apoio Potencia CADocumento19 pagineUninter - Caderno de Apoio Potencia CAGil XavierNessuna valutazione finora
- Hfen10 Teste Final 2 ResolucaoDocumento1 paginaHfen10 Teste Final 2 Resolucaofilipa50% (2)
- Chave MatheusDocumento4 pagineChave MatheusFernando Borges QueirozNessuna valutazione finora
- Motores InduçãoDocumento3 pagineMotores InduçãoMariana AndradeNessuna valutazione finora
- Equilíbrio EletroquímicaDocumento26 pagineEquilíbrio EletroquímicaHeloiseNessuna valutazione finora
- Orçamento Ultralar Mudanças Aproveitamento de Carga - SR Cleber - São Paulo SP X Olinda PeDocumento3 pagineOrçamento Ultralar Mudanças Aproveitamento de Carga - SR Cleber - São Paulo SP X Olinda PeCleber RodriguesNessuna valutazione finora
- Manual Guia FR 10-02-22 SiteDocumento2 pagineManual Guia FR 10-02-22 SiteRodrigo ZanettiNessuna valutazione finora
- AgitadoresDocumento2 pagineAgitadoresHugo Charles KlugerNessuna valutazione finora
- Manual Torno PRN-320 IMORDocumento34 pagineManual Torno PRN-320 IMORRenato Bednarski Ramos82% (17)
- 3852Documento40 pagine3852RODRIGO_RALONessuna valutazione finora
- Declaracao CLCB 2344724Documento2 pagineDeclaracao CLCB 2344724popyzmanNessuna valutazione finora
- Recuperação SecundáriaDocumento12 pagineRecuperação SecundáriaDélcio Dos SantosNessuna valutazione finora
- Apostila DispEletricoDocumento7 pagineApostila DispEletricotokafita100% (4)
- Treinamento piloto comercial aviãoDocumento3 pagineTreinamento piloto comercial aviãoReinaldy Rodrigues Rocha DiNessuna valutazione finora
- Conceitos básicos de telecomunicações para iniciantesDocumento33 pagineConceitos básicos de telecomunicações para iniciantesRafaelRoessler100% (3)
- Resolucao Da Lista de Exercicios 3 - Calculos Estequiometricos - Parte 2 - 1 Bimestre 2013 - 2 SeriesDocumento4 pagineResolucao Da Lista de Exercicios 3 - Calculos Estequiometricos - Parte 2 - 1 Bimestre 2013 - 2 SeriesRafael CostaNessuna valutazione finora
- Barragem Dorta Gs TCC IlhaDocumento87 pagineBarragem Dorta Gs TCC IlhaMário FailacheNessuna valutazione finora
- Introdução aos conceitos básicos de físicaDocumento126 pagineIntrodução aos conceitos básicos de físicacutiacelNessuna valutazione finora
- Introdução ao estudo da tecnologia de alimentosDocumento11 pagineIntrodução ao estudo da tecnologia de alimentosFabiane FariasNessuna valutazione finora
- Refrigeração ParcialDocumento3 pagineRefrigeração ParcialRenato FilhoNessuna valutazione finora
- Catalogo Produto Rtac (RLC Prc019f PB)Documento49 pagineCatalogo Produto Rtac (RLC Prc019f PB)Rodrigo SantosNessuna valutazione finora
- Leiautes Da EFD-Reinf v1.4 - Anexo I - TabelasDocumento65 pagineLeiautes Da EFD-Reinf v1.4 - Anexo I - TabelasSidnei JosephikNessuna valutazione finora
- Esquema Eletrico Iveco Vertis Euro 5 HD 90V18 e HD 130V19Documento35 pagineEsquema Eletrico Iveco Vertis Euro 5 HD 90V18 e HD 130V19Alberto Matties88% (8)
- Refrigerador de Cerveja DidáticoDocumento30 pagineRefrigerador de Cerveja DidáticoLurriane PereiraNessuna valutazione finora
- Equipamentos para Trabalho em AlturaDocumento17 pagineEquipamentos para Trabalho em AlturaAlessandro Antunes de AmorimNessuna valutazione finora
- Parecer Analítico Sobre Regras Regulatórias - SEAE PDFDocumento18 pagineParecer Analítico Sobre Regras Regulatórias - SEAE PDFbrunofvieiraNessuna valutazione finora
- Isolamento térmico e impermeabilização de coberturas em terraçoDocumento91 pagineIsolamento térmico e impermeabilização de coberturas em terraçoSantos Cesar Dos Santos César CesarNessuna valutazione finora
- Plano de Negócios Do Pólo Da TilapiaDocumento83 paginePlano de Negócios Do Pólo Da TilapiaCláudio Silva50% (2)