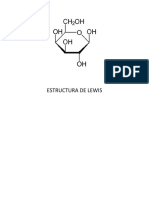
Potrebbero piacerti anche
- Reporte 1 Digestión de MacromoleculasDocumento6 pagineReporte 1 Digestión de MacromoleculasCarol CacerosNessuna valutazione finora
- Determinacion Parametros Vmax y KM LastDocumento7 pagineDeterminacion Parametros Vmax y KM LastJonatan Ramirez MercadoNessuna valutazione finora
- La InvertasaDocumento2 pagineLa InvertasaDavid Saúl Paredes Puma100% (2)
- Tipos de Vectores de Clonación y ExpresiónDocumento9 pagineTipos de Vectores de Clonación y ExpresiónMarcia100% (2)
- Biorreactor AirliftDocumento60 pagineBiorreactor AirliftAlexis GonzalezNessuna valutazione finora
- Prácticas de Frutas y Hort. 1,2 y 3.Documento22 paginePrácticas de Frutas y Hort. 1,2 y 3.Grupo 4 AgroindustriasNessuna valutazione finora
- Modelo de MonodDocumento5 pagineModelo de MonodyenyNessuna valutazione finora
- Problemas Formulacion y CineticaDocumento2 pagineProblemas Formulacion y CineticaAbryNessuna valutazione finora
- Determinacion de Humedad y Ceniza, Diego VargasDocumento2 pagineDeterminacion de Humedad y Ceniza, Diego VargasCorin Daniela NarvaezNessuna valutazione finora
- Ejercicios Cinética Michaelis MentenDocumento2 pagineEjercicios Cinética Michaelis MentenDavid Esteban Soler Camargo100% (1)
- Ecuación de Michaelis-Menten (Deducción)Documento2 pagineEcuación de Michaelis-Menten (Deducción)Eduardo CienfuegosNessuna valutazione finora
- Balance de Biorreactor Por Lote AlimentadoDocumento1 paginaBalance de Biorreactor Por Lote AlimentadoLuz RoldanNessuna valutazione finora
- Tema 4 Farmacocinética 2021-2Documento116 pagineTema 4 Farmacocinética 2021-2Athenas A.P100% (1)
- APLICACIÓN KLaDocumento2 pagineAPLICACIÓN KLaClau BecerrilNessuna valutazione finora
- Problemas Cinetica Propuestos para El ExamenDocumento3 pagineProblemas Cinetica Propuestos para El ExamenBlas Alfaro StefanyNessuna valutazione finora
- Tarea 3Documento2 pagineTarea 3Kagney UralesNessuna valutazione finora
- 2 Extracción de Enzima Lipasa y Medición de La Actividad EnzimáticaDocumento5 pagine2 Extracción de Enzima Lipasa y Medición de La Actividad Enzimáticajuan esteban perezNessuna valutazione finora
- Problema - ExamenDocumento2 pagineProblema - ExamenAlejandra Cuzco100% (3)
- 3 Clase de Problemas Actividad Enzimas 1Documento5 pagine3 Clase de Problemas Actividad Enzimas 1Julio Cesar Castañeda Mancipe80% (5)
- Extracción de Invertasa PDFDocumento3 pagineExtracción de Invertasa PDFRonald André Buleje HinostrozaNessuna valutazione finora
- Respuestas Taller Problemas Internet PDFDocumento13 pagineRespuestas Taller Problemas Internet PDFMagda CastillaNessuna valutazione finora
- Cultivo ContinuoDocumento13 pagineCultivo ContinuoCatherine AntonietaNessuna valutazione finora
- Extracción EnzimáticaDocumento9 pagineExtracción EnzimáticaGuadalupe MunizNessuna valutazione finora
- MediosDocumento6 pagineMediosDenisse MotaNessuna valutazione finora
- Transferencia de Oxigeno en BioreactoresDocumento45 pagineTransferencia de Oxigeno en BioreactoresCarlos Coronado LezmaNessuna valutazione finora
- Cinetica Enzimatica de La Invertasa Mediante La Determinacion de La Velocidad Inicial de ReacciãnDocumento9 pagineCinetica Enzimatica de La Invertasa Mediante La Determinacion de La Velocidad Inicial de Reacciãnnicolas fetecuaNessuna valutazione finora
- Crecimiento BacterianoDocumento5 pagineCrecimiento Bacterianodavid espinoza avendaño67% (3)
- 4efecto de La Actividad Enzimática de Inulinasas Usando Metales MG y Co Como Posibles Activador o Inhibidor Utilizando Kluyveromyces MarxianusDocumento8 pagine4efecto de La Actividad Enzimática de Inulinasas Usando Metales MG y Co Como Posibles Activador o Inhibidor Utilizando Kluyveromyces MarxianusEnrique GomezNessuna valutazione finora
- Tercer Parcial Ingeniería EnzimáticaDocumento6 pagineTercer Parcial Ingeniería EnzimáticaSalma PeñalOveeNessuna valutazione finora
- Practica 5 LIIBIDocumento33 paginePractica 5 LIIBICésar Porras100% (1)
- Problemas Cinetica EnzimaticaDocumento9 pagineProblemas Cinetica EnzimaticamichypaoNessuna valutazione finora
- GalactosaDocumento9 pagineGalactosaGUTIÉRREZ SÁNCHEZ OSCARNessuna valutazione finora
- Prueba Rojo Fenol Campana DurhamDocumento7 paginePrueba Rojo Fenol Campana DurhamAlejandra GualdronNessuna valutazione finora
- Ejercicios Cinética 2Documento3 pagineEjercicios Cinética 2Mayra MoralesNessuna valutazione finora
- Escalamiento de Un BioprocesoDocumento16 pagineEscalamiento de Un BioprocesoYaneli Nicole Champi CorcueraNessuna valutazione finora
- Parte 1:: Cinética de Crecimiento MicrobianoDocumento72 pagineParte 1:: Cinética de Crecimiento MicrobianoYandira Gutierrez MendozaNessuna valutazione finora
- Producción de Penicilina G A Partir de Penicilium ChrysogenumDocumento13 pagineProducción de Penicilina G A Partir de Penicilium ChrysogenumAlejandra Rubiano0% (1)
- Utlizacion de Carbohidratos y Acidos OrganicosDocumento46 pagineUtlizacion de Carbohidratos y Acidos OrganicosRoberto PopocaNessuna valutazione finora
- Medición de Coeficientes de Transferencia de Calor en Diferentes GeometriasDocumento6 pagineMedición de Coeficientes de Transferencia de Calor en Diferentes Geometriasjessicasanchez-314Nessuna valutazione finora
- Ejercicios KLa Coeficiente de Transferencia de MasaDocumento12 pagineEjercicios KLa Coeficiente de Transferencia de MasaGabriela Kaulitz Jahseh67% (3)
- Taller EnzimasDocumento5 pagineTaller Enzimaswilber menesesNessuna valutazione finora
- Digestion de CarbohidratosDocumento5 pagineDigestion de CarbohidratosEvelyn Jim 32950% (1)
- Guía de Problemas para Cultivo Continuo y AlimentadoDocumento3 pagineGuía de Problemas para Cultivo Continuo y AlimentadoWambaNessuna valutazione finora
- Ejercicios Cultivo en Lote para EstudiantesDocumento1 paginaEjercicios Cultivo en Lote para EstudiantesBarbie Centeno33% (3)
- Cuantificación de Proteina de Clara de HuevoDocumento1 paginaCuantificación de Proteina de Clara de HuevoAbel de Jesús RiveraNessuna valutazione finora
- Libro de Cinetica MicrobianaDocumento24 pagineLibro de Cinetica MicrobianaCristian Danilo Hernandez FigueroaNessuna valutazione finora
- Eq 7 Microbiologia IndustrialDocumento101 pagineEq 7 Microbiologia IndustrialAdelheint Bernstein75% (4)
- Bioquímica Practica 4Documento8 pagineBioquímica Practica 4Andres Fernando Silvestre SuarezNessuna valutazione finora
- Cuestionario EnzimasDocumento5 pagineCuestionario EnzimasAmelia GómezNessuna valutazione finora
- Facultad de Agronomía: Licenciatura en Agronomía Bioquímica Coria Gil Norma Ana BeliDocumento9 pagineFacultad de Agronomía: Licenciatura en Agronomía Bioquímica Coria Gil Norma Ana BeliFREDDY SOBERANONessuna valutazione finora
- Biotransformación Fase I y II de Compuestos XenobiDocumento4 pagineBiotransformación Fase I y II de Compuestos XenobiAntonio Tenorio GonzalezNessuna valutazione finora
- 6 y 7 Inf. 1Documento21 pagine6 y 7 Inf. 1Yoselin Jazmin Azabache AbdhalaNessuna valutazione finora
- Metabolismo de XenobioticosDocumento4 pagineMetabolismo de XenobioticosGerson SanchezNessuna valutazione finora
- Glucosa OxidasaDocumento9 pagineGlucosa OxidasaGermán Gamio Farfán0% (1)
- Reporte 1, Sobre EnzimasDocumento5 pagineReporte 1, Sobre EnzimasIsabella JiménezNessuna valutazione finora
- Hidrólisis de GlucósidosDocumento10 pagineHidrólisis de GlucósidosAlberth PedemonteNessuna valutazione finora
- Enzimas en Los AlimentosDocumento65 pagineEnzimas en Los Alimentosluis gustavorNessuna valutazione finora
- Enzimas en AlimentosDocumento25 pagineEnzimas en AlimentosJR Bio GenNessuna valutazione finora
- Bioquimica - 57 - Enzimologia - y - Bioenergetica - Nestor Yeudy Perea GamboaDocumento7 pagineBioquimica - 57 - Enzimologia - y - Bioenergetica - Nestor Yeudy Perea GamboaNestor Perea GamboaNessuna valutazione finora
- Discusiones Sobre Acido Ascorbico Tesis PDFDocumento14 pagineDiscusiones Sobre Acido Ascorbico Tesis PDFRandy Omar BecerraNessuna valutazione finora
- Isotermas de SorciónDocumento15 pagineIsotermas de SorciónGladys'Ameriica PraadooNessuna valutazione finora
- Capitulo 9 Cromatografía de Gases (Traduccion)Documento44 pagineCapitulo 9 Cromatografía de Gases (Traduccion)Gladys'Ameriica PraadooNessuna valutazione finora
- Reporte Cuantificación de ProteínasDocumento16 pagineReporte Cuantificación de ProteínasGladys'Ameriica PraadooNessuna valutazione finora
- Análisis SensorialDocumento41 pagineAnálisis SensorialGladys'Ameriica PraadooNessuna valutazione finora
- Geles de AlmidónDocumento6 pagineGeles de AlmidónGladys'Ameriica PraadooNessuna valutazione finora
- Tratamiento de Aguas Con LirioDocumento6 pagineTratamiento de Aguas Con LirioGladys'Ameriica PraadooNessuna valutazione finora
- Lirio Acuático AprovechamientoDocumento6 pagineLirio Acuático AprovechamientoGladys'Ameriica PraadooNessuna valutazione finora
- Gomas Aplicadas en El Área de AlimentosDocumento21 pagineGomas Aplicadas en El Área de AlimentosGladys'Ameriica PraadooNessuna valutazione finora
- Fisica MCUDocumento9 pagineFisica MCURobertNessuna valutazione finora
- Guia 4 - Separación de Mezclas Homogéneas y Heterogéneas-Signed-SignedDocumento3 pagineGuia 4 - Separación de Mezclas Homogéneas y Heterogéneas-Signed-Signeddiego boneNessuna valutazione finora
- Electrodeposicion Del OroDocumento6 pagineElectrodeposicion Del OroMelissa LP50% (6)
- 02 - Física en Procesos Industriales - Control V1Documento3 pagine02 - Física en Procesos Industriales - Control V1Ricardo Muñoz SuazoNessuna valutazione finora
- SUSTANCIAS PURAS Unidad 3 Termofluidos CDocumento32 pagineSUSTANCIAS PURAS Unidad 3 Termofluidos CJose VenegasNessuna valutazione finora
- Conferencia 1Documento8 pagineConferencia 1Denis Lopez MartinezNessuna valutazione finora
- Tema 6 Estabilidad de TaludDocumento58 pagineTema 6 Estabilidad de TaludCarlos EscobarNessuna valutazione finora
- Analisis Instrumentalde Alimentos EDUNIV2021Documento538 pagineAnalisis Instrumentalde Alimentos EDUNIV2021danNessuna valutazione finora
- FILTRO TAMBOR Practica Tec LagunaDocumento12 pagineFILTRO TAMBOR Practica Tec LagunaBrenda Nollely ReyesNessuna valutazione finora
- Batimetria Lago de Tota PDFDocumento40 pagineBatimetria Lago de Tota PDFLau VhNessuna valutazione finora
- Julio César Vela VelazquezDocumento12 pagineJulio César Vela VelazquezCarlo Enrique Hernandez Rojas QuijadaNessuna valutazione finora
- Informe Viscocidad Metodo de StokesDocumento9 pagineInforme Viscocidad Metodo de StokesAntonio Cortez QuispeNessuna valutazione finora
- Actividad 7. Sustancia PuraDocumento4 pagineActividad 7. Sustancia PuraMichelle Boesler100% (1)
- Sales-Oxisales-organizador VisualDocumento4 pagineSales-Oxisales-organizador VisualAyde CachicatariNessuna valutazione finora
- Qué Es El Rombo NFPADocumento4 pagineQué Es El Rombo NFPAsoraya barretoNessuna valutazione finora
- Termodinamica UnadDocumento14 pagineTermodinamica UnadDarwin Andres Beleño GuillenNessuna valutazione finora
- Movimientos PlanetariosDocumento19 pagineMovimientos PlanetariosJaniel Paredes MoralesNessuna valutazione finora
- Determinacion de HAP's en Aire AmbientalDocumento8 pagineDeterminacion de HAP's en Aire Ambientalsupervisorlab ihaNessuna valutazione finora
- Taller de Química y SaludDocumento1 paginaTaller de Química y SaludZerímar AblemNessuna valutazione finora
- Decoloración Fenolftaleína en Medio BásicoDocumento7 pagineDecoloración Fenolftaleína en Medio BásicoDaniel Castillo hernándezNessuna valutazione finora
- 2001 Tesis Y.aznarDocumento277 pagine2001 Tesis Y.aznarCristian Felipe ZapataNessuna valutazione finora
- Guia de Laboratorio IV. Propiedades e Identificación de Los Carbohidratos Quimica OrganicaDocumento4 pagineGuia de Laboratorio IV. Propiedades e Identificación de Los Carbohidratos Quimica OrganicaMayerlinNessuna valutazione finora
- Investigación Sobre La EnergíaDocumento6 pagineInvestigación Sobre La EnergíaCamila SorianoNessuna valutazione finora
- Grupo2 InformedesulfatosDocumento56 pagineGrupo2 InformedesulfatosKennedy Marquez MuñozNessuna valutazione finora
- 8501-15 Fisicoquimica WordDocumento106 pagine8501-15 Fisicoquimica WordMariannaGarciaSalazarNessuna valutazione finora
- Equivalente GramoDocumento6 pagineEquivalente GramoMarina AquinoNessuna valutazione finora
- Errores Cometidos en La Medicion Con CintaDocumento9 pagineErrores Cometidos en La Medicion Con CintaJULIAN HERNANDO SEGURA LAGUNANessuna valutazione finora
- Apunte EXPLORACIÓN Estudios GeoquímicosDocumento16 pagineApunte EXPLORACIÓN Estudios GeoquímicosDaniela Olivares SedanNessuna valutazione finora
- Factores Que Influyen en La Eficacia deDocumento2 pagineFactores Que Influyen en La Eficacia deThalia Cóndor FloresNessuna valutazione finora
- Hoja de Datos Motor Trifasico w22 13415827Documento1 paginaHoja de Datos Motor Trifasico w22 13415827Carlos TovarNessuna valutazione finora