Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
Resumen Completo Anatomia II
Caricato da
sofiaCopyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
Formati disponibili
Resumen Completo Anatomia II
Caricato da
sofiaCopyright:
Formati disponibili
lOMoARcPSD|1684066
Resumen completo Anatomia II
Neuroanatomía (Universidade de Santiago de Compostela)
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Para comenzar con el estudio del sistema nervioso podemos hacer una primera
clasificación, muy general, que es la siguiente:
• Sistema Nervioso Central (SNC): es el sistema nervioso situado dentro de la
cavidad craneal o dentro del conducto raquídeo, de forma que está siempre
dentro de alguna estructura de protección.
• Sistema Nervioso Periférico (SNP): se trata de una serie de nervios, que se
denominan periféricos y que nacen en el SNC. Estos nervios periféricos llegan a
todas las estructuras de nuestro organismo.
Cabe decir también que el sistema nervioso central está tremendamente protegido
por una serie de estructuras, que enumeramos a continuación:
• Las meninges: son las primeras envolturas de protección del sistema nervioso
central. Se trata de una especie de “telillas” que envuelven las estructuras
nerviosas y las protegen del entorno.
• Líquido cefalorraquídeo: se trata de un fluido especial que es segregado por
vasos y que se encuentra alrededor del sistema nervioso y también incluido
dentro de ciertas cavidades y conductos del SNC. Este líquido, por el principio
de Arquímedes ejerce una fuerza que mantiene al sistema nervioso en su
posición sin ejercer presión sobre él.
• Cavidad craneal y conducto raquídeo: se trata de estructuras de protección de
estirpe ósea cuya principal función es evitar traumas en el SNC, e impedir así
que sea dañado.
OBSERVACIÓN DE ESTRUCTURAS DEL SNC EN LA SALA DE DISECCIÓN
Para poder obervar las distintas estructuras del SNC en la sala de disección,
emplearemos dos técnicas diferentes, cuyo uso dependera de aquellas estructuras
específicas que deseemos observar:
• Vía auricular: se trata de realizar una incisión de oreja a oreja que permita
extraer la piel y el tejido celular subcutáneo, para poder observar así la calota
craneal. Posteriormente se trepana el cráneo y observamos las superficies
meníngeas. Esta técnica permite la observación del encéfalo.
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Incisión dorsal media: esta técnica se emplea para la observación de la médula
espinal y consiste en realizar una incisión larga en la región medio-dorsal del
cuerpo, poniendo de manifiesto las vértebras.
CLASIFICACIÓN DEL SISTEMA NERVIOSO CENTRAL
El sistema nervioso central lo clasificamos según el lugar donde se sitúe, de forma
que podemos diferenciar las siguientes estructuras:
a) Encéfalo: aquella parte del SNC situado dentro del cráneo. A su vez, el encéfalo
está dividido en varias partes:
a. Cerebro: este a su vez estará dividido en dos nuevas partes:
i. Telencéfalo (parte más externa)
ii. Diencéfalo (situada en el centro, casi no se ve nada por fuera,
sólo un poco por la cara basal)
b. Cerebro pequeño o cerebelo: es un órgano único. Está unido con el
tronco del encéfalo mediante los pedúnculos cerebelosos (que son fibras
de sustancia blanca). Existen tres pares de pedúnculos cerebelosos que se
denominan:
i. Pedúnculos cerebelosos inferior.
ii. Pedúnculos cerebelosos medios.
iii. Pedúnculos cerebelosos inferiores.
c. Tronco del encéfalo: en él distinguimos tres partes o “pisos”:
i. Mesencéfalo (parte superior)
ii. Puente (parte media)
iii. Bulbo (parte inferior)
b) Médula espinal: la parte del SNC situada dentro del conducto raquídeo.
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CORTES DEL SISTEMA NERVIOSO CENTRAL
En todos los cortes de Sistema Nervioso Central observamos tres elementos
totalmente claros: la materia o sustancia blanca, la materia o sustancia gris y las
diferentes cavidades y conductos que rodean y atraviesan el SNC.
El sistema nervioso está compuesto por muchas células, que son las neuronas,
así como otras células que permiten que se mantengan vivas las neuronas (son las
células de la glía). Las neuronas se caracterizan por tener un cuerpo o soma neuronal y
unas prolongaciones (dendritas y axones), de forma que la neurona puede enviar
información de unos sitios a otros. Cuando muchos somas neuronales se agrupan juntos,
dan lugar a lo que se conoce como sustancia gris (son acúmulos de cuerpos neuronales).
La sustancia blanca es el resultado de la acumulación de prolongaciones neuronales
(normalmente axones).
Cabe comentar el hecho de que en la sustancia blanca, si aparece algún soma
neuronal en los cortes histológicos, pero no se “apiñan” como ocurre en la sustancia
gris. De esta forma también aparecen fibras nerviosas no agrupadas en la sustancia gris.
La sustancia gris, en el SNC a veces se sitúa en la superficie, mientras que en otras
ocasiones se sitúan más en profundidad, formando unas pequeñas agrupaciones que se
denominan núcleos.
En cuanto a las cavidades, es preciso destacar que el cerebro tiene una serie de
ellas que se denominan ventrículos, y que están comunicados con los otros ventrículos y
con el exterior mediante una serie de conductos. Tanto ventrículos como conductos van
a estar ocupados siempre por el LCR.
LÍMITES DE LAS DISTINTAS PARTES DEL SNC
Los límites entre los componentes del SNC pueden ser “naturales” o por
convención. Esto se debe a que existen órganos que están muy bien delimitados,
mientras que otras estructuras presentan unos límites, cuanto menos, indefinidos. En
estos casos es necesario tomar un punto anatómico que permita definir dónde comienza
un determinado órgano y donde termina el otro. Algunos de los límites son los que a
continuación presentamos:
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Límite entre la médula espinal y el bulbo raquídeo: se emplea una
“interrupción” en el surco medio anterior de bulbo y médula que se
denomina Decusación motriz o de Misticheli. Esta decusación es el
resultado del cruzamiento de fibras que se encargan del movimiento
voluntario. En el lugar donde se cruzan, se traza un plano, y lo que está
por encima de dicho plano se denomina bulbo y lo que está por debajo,
médula. Existe una pequeña parte del bulbo que se encuentra en el
conducto raquídeo (queda por debajo del agujero magno), de forma que
la decusación motriz se sitúa a nivel de las articulaciones occípito-
atloideas.
• Límites de la Protuberancia o Puente de Varolio: el puente sobresale
mucho del tronco del encéfalo, de forma que existen dos surcos, uno por
arriba y otro por abajo, que se denominan surco póntico superior (o surco
ponto-mesencefálico) y surco póntico inferior (o surco bulbopóntico), y
que permiten establecer fácilmente los límites de la protuberancia.
• Límite superior del tronco del encéfalo: tras el quiasma óptico sale la
cintilla o bandeleta óptica que rodea el tronco del encéfalo. Así, las vías
ópticas se consideran el límite superior del tronco del encéfalo. La zona
que queda en el medio, es una zona diencefálica que se introduce ahí a
modo de cuña. Esto provocaría la formación de unos límites complejos,
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
pero éstos se simplifican. El triángulo diencefálico resultante se
encuentra entre dos pedúnculos cerebrales y la cintilla óptico, y por ello
se denomina triángulo interpeduncular u opto-peduncular.
mesencéfal
Es una cuña del
o
diencéfalo, la única Surco póntico
parte del superior.
diencéfalo triángulo opto- Surco
que vemos por peduncular pontomesencefálico
fuera agujero Surco póntico
ciego inferior.
Surco bulbopóntico.
Surco medio anterior del
decusación bulbo
Surco medio anterior de la médula
motriz espinal.
NEURONAS Y CÉLULAS GLIALES
Lo primero que llama la atención es que el SNC, es que se trata de una estructura
diferente al resto de los órganos, pues no posee una estructura sólida, sino que desde el
punto de vista estructural es una estructura semisólida o casi gelatinosa. Tanto es así que
no se puede suturar el SN con aguja e hilo (salvo la parte externa, las meninges). Este
aspecto gelatinoso se debe a la pequeñísima cantidad de tejido conjuntivo que existe en
el SNC, dado que el único tejido conjuntivo presente es el que aportan los vasos.
De esta manera, el SNC va a estar compuesto sólo por dos tipos de células: las
neuronas y las células de la glía. Las neuronas son las células nobles del SNC y las
células de la glía son imprescindibles para la vitalidad de las neuronas. De hecho hay
diez veces más células de la glía que neuronas.
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
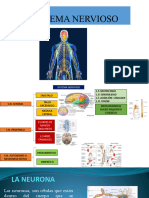
LAS NEURONAS
Se calcula que el SN humano puede tener una aproximación de 100.000 millones
de neuronas. Lo que ocurre, es que estas neuronas son tan especializadas que no tienen
capacidad de división, por lo que nosotros nacemos con un número determinado de
neuronas que no aumentará, y que llamamos “patrimonio neuronal”. Lo único que
podemos hacer a lo largo de la vida es dilapidar esas neuronas, y a partir de los treinta
años se pierden unas cien mil neuronas diarias. A esto se le conoce como pérdida
fisiológica de neuronas. Pero esta cantidad puede verse aumentada por distintos
factores: elementos tóxicos, usos diversos del sistema nervioso….
No obstante, cabe señalar también que existen unas células madre en el sistema
nervioso que se sitúan normalmente pegadas a las zonas ventriculares, así como también
ciertas localizaciones cerebrales en las que existen neuronas capaces de regenerarse.
Esto sucede en el lóbulo olfatorio, donde las neuronas se regeneran mensualmente.
En cuanto a las características estructurales, cabe decir que las neuronas,
como cualquier célula posee un núcleo inmerso en un cuerpo celular (que se denomina
soma o pericárion). Este soma neuronal puede ser de muchas formas, de hecho las
neuronas se clasifican en función de su morfología: fusiformes, piriformes,
estrelladas… El soma está limitado por una membrana plasmática, pero que posee
algunas características propias. Esto se debe a que la neurona necesita mucha superficie
de membrana para establecer sinapsis con otras neuronas. Para aumentar la superficie de
la neurona, de la membrana salen unas prolongaciones en todas direcciones. Estas
prolongaciones reciben el nombre de dendritas, y desde el punto de vista estructural son
similares al cuerpo de la neurona, pero son más alargadas para permitir la existencia de
más contactos con otras neuronas, dado que el número medio de sinapsis de cada
neurona es de 14.000 (aunque hay algunas en las que la cifra alcanza las 100.000). Por
ello hay también muchas neuronas que poseen espinas dendríticas, localizaciones
específicas en las que tienen lugar las sinapsis. Esto se ha demostrado en estudios en los
que, privando a animales de luz desde el nacimiento, no se desarrollaron espinas
dendríticas en núcleos talámicos relacionados con la visión. Esto demuestra dos cosas:
en primer lugar, que las espinas dendríticas están en relación con los contactos
sinápticos, y en segundo lugar, que en las neuronas pueden aumentar o disminuir sus
contactos sinápticos. Esto está muy acorde con lo que se conoce como “Ley de Wolf”,
que afirma que la función hace al órgano. Pero hay que especificar: las neuronas no
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
aumentan, pero las sinapsis se hacen más eficientes porque aumenta sensiblemente el
número de ellas.
En las neuronas existen diferencias en el número de dendritas, lo que permite
clasificarlas atendiendo a este aspecto en multipolares, bipolares, monopolares… Cabe
decir también, que las dendritas del vértice de las neuronas se denominan apicales, y las
que están en la base, basales.
Las neuronas deben conectarse unas con otras, y algunas se encuentran muy
lejanas (incluso a más de un metro de distancia). Las prolongaciones largas que
permiten estos contactos reciben el nombre de axones. Existe un único axón por
neurona, que puede salir del cuerpo de la neurona o de la base de las dendritas. Parten
de una zona que se semeja a un embudo y que se denomina cono axónico. A partir de
ahí, el axón tendrá el mismo grosor a lo largo de todo el recorrido (sólo cambia en el
teleaxón), mientras que las dendritas son más gruesas cuanto más cerca estén del soma
neuronal. Una particularidad llamativa, es que las prolongaciones del axón parten
siempre de forma perpendicular, al contrario de lo que pasaba con las dendritas, cuyas
prolongaciones partían en forma irregular dando un aspecto “árboreo” al conjunto.
Uno de los descubrimientos de Cajal, que anunció junto con la teoría de la
neurona, fue la Ley de polarización dinámica (que se cumple en el 99% de los casos)
que sostiene que la neurona tiene dos polos:
- Polo aferente (sensitivo): recibe las órdenes. Lo constituyen el cuerpo de
la neurona y las dendritas.
- Polo eferente (motor): polo que manda órdenes o lleva información a
otros emplazamientos. El polo eferente lo constituye el axón.
De esta manera, la neurona recibe la información por soma y dendritas, y la
envía a través del axón.
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El sistema nervioso integra todas las informaciones que le llegan (internas y
externas) y da respuestas globales. Así, el SNC es el motor de todo, y lo fundamental
de él es que hace reaccionar al cuerpo como una unidad completa. El sistema más
simple es la neurona, que recibe, integra y transmite la información. De ahí que cuantas
más neuronas existan en el sistema nervioso, mayor será su complejidad.
En organismos simples el SN puede estar compuesto por una única neurona, que recibe,
procesa y transmite la información, pero el sistema puede complicarse. En estos casos,
las neuronas empiezan a especializarse. Algunas se especializan en recibir información
y otras en dar respuestas. Así se forman complejos de neuronas aferentes sensitivas y
neuronas eferentes motoras.
Aún así, cuando los sistemas nerviosos adquieren una mayor complejidad, entre
las neuronas sensitivas y las motoras, se sitúa un tercer tipo: las interneuronas o
neuronas de asociación. En relación con la cantidad de interneuronas que tengamos,
daremos unas respuestas más o menos elaboradas. Por ejemplo, en el sistema nervioso
humano el 99% de las neuronas son interneuronas, lo que hace que podamos pensar,
hablar, tener sentimientos frente a congéneres, dar respuestas totalmente diferentes ante
los mismos estímulos, etc.
Es muy sencillo explorar las neuronas sensitivas y las motoras, mientras que
examinar las interneuronas es tremendamente difícil, tarea en lo que ayuda
especialmente la psicología.
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Otro aspecto a destacar sobre las neuronas, es que deben poder mantener vivas
sus prolongaciones. Esto se realiza mediante el “transporte axónico o axoplásmico” en
el que el cuerpo de la neurona tiene que enviar sustancias a la parte final del axón y
recibir sustancias de deshecho procedentes del mismo. Este transporte puede ser
anterógrado o retrógado. Por el transporte anterógrado, se envían sustancias desde el
cuerpo de la neurona hasta el final del axón y está mediado por una proteína específica:
la cinesina. El transporte retrógrado va desde los axones hasta el cuerpo de la neurona,
y está mediado por la dineína.
El transporte axónico puede ser de dos tipos: rápido (de medio metro por cada
24 h) y lento (5 mm por cada 24 h). Este transporte ha servido para que los
neuroanatómicos pudieran saber hasta donde llegan las neuronas, mediante el empleo de
ciertas sustancias marcadoras que penetran por el axón y llegan hasta el soma neuronal.
El núcleo que podemos observar en la neurona presenta un nucleolo muy
marcado. Rodeando al núcleo, en la zona del citoplasma tienen todas las organelas que
poseen todas las células, pero algunas son más manifiestas.
Muchas de ellas tienen un retículo endoplásmico rugoso enormemente
abundante, pues la neurona necesita gran cantidad de proteínas (neurotransmisores,
neuromoduladores,….). Lo mismo sucede con el aparato de Golgi, que también estará
muy desarrollado. El retículo endoplasmático rugoso de las neuronas reciben un nombre
específico: Sustancia de Nissl (Sustancia Tigroide de Nissl), que ya que posee ácidos
ribonucleicos, se tiñe con colorantes básicos (específicamente con el violeta de cresilo o
el azul de toluidina, llamada también Tinción de Nissl). Si no observamos la sustancia
tigroide, podemos decir que la neurona estaba estimulada en el momento de la fijación,
mientras que si aparece quiere decir que la neurona se encuentraba en reposo.
En el cuerpo también existen mitocondrias, bastante abundantes, que se usan
para el mecanismo oxidativo, pero con una característica particular: únicamente utilizan
como nutrientes la glucosa y, obviamente, el oxígeno. Así cuando una neurona o grupo
de neuronas se quedan sin alguno de sus nutrientes básicos puede producirse la muerte
neuronal. De ahí que el cerebro vaya a consumir aproximadamente el 20 por ciento de
todo el oxígeno del organismo. Lo mismo ocurre con la glucosa, hasta el punto de que
cuando baja la glucosa en sangre el paciente puede entrar en coma.
Existen también neuronas que acumulan sustancias de deshecho, sobre todo
lípidos. Algunas se caracterizan por adquirir una coloración parda en función de lo que
acumulen en su interior.
10
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Cuando hacemos un estudio histológico del SN, podemos observar el cuerpo y
algunas prolongaciones neuronales, pero podemos hacer también un estudio para poder
ver una neurona con todas sus prolongaciones. Esto se realiza con la coloración de
Golgi, pero se trata de una técnica complicada, porque no tiñe todas las neuronas sino
únicamente algunas. Por ello la técnica requiere una cierta habilidad por parte del
histólogo. Esta tinción fue especialmente importante para Cajal, pues le sirvió para
demostrar la mayor parte de sus trabajos.
También pueden hacerse tinciones específicas, por ejemplo para marcar las
microfibrillas (microtúbulos y microfilamentos), como sería la plata reducida de Cajal.
CÉLULAS DE LA GLÍA
Dado que el SNC carece de tejido conjuntivo, para su supervivencia son necesarias
las células de la Glía. Actualmente, sigue en vigor la clasificación realizada por Del Río
Hortega, que dividía la Glía en dos tipos:
a) Macroglía: puede existir tanto en el SNC como en el SNP, y se divide en dos
grandes grupos:
a. Astrocitos: aparecen en el SNC como tal y en el SNP como células
satélites de los ganglios (al lado de las neuronas).
b. Oligodendrocitos: como oligodendrocitos los vemos sólo en el SNC, ya
que en el SNP reciben el nombre de células de Schwann.
b) Microglía: las células que componen esta división fueron consideradas parte de
la glía, pero en realidad no pueden ser consideradas como células gliales, ya que
se trata de células de extirpe conjuntiva y forman parte del sistema de defensa
del organismo (son macrófagos).
Cabe decir que en la actualidad, a esta clasificación se le han añadido las células
ependimarias.
LOS ASTROCITOS
Estas células, como toda la glía, son ectodérmicas (el tejido conjuntivo es
mesodérmico). Se denominan astrocitos porque tienen una forma estrellada
característica (cuerpo y prolongaciones), cuyas prolongaciones llegan hasta las
11
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
neuronasny entran en contacto con ellas. Del mismo modo, dichas prolongaciones
llegan también hasta los vasos, de forma que éstos quedan cubiertos por pies
astrocitarios, formando lo que se conoce como barrera hematoencefálica. Finalmente
otros astrocitos se dirigen hacia la superficie externa del sistema nervioso, donde se
encuentra el límite con las cavidades que contienen el LCR.
Gracias a las características de los astrositos, todo lo que llega hasta la neurona va a
estar controlado por ellos. De ahí que los clásicos denominaran los pies astrocitarios
como “pies chupadores”, pues hacen una función de “filtro” para lo que llega a la
neurona.
Las sustancias pueden pasar a través de los astrocitos o a través de los pequeños
orificios que éstos dejan entre sus pies. Lo mismo ocurre con las sustancias de
deshecho.
Hay estudiosos que consideran que los astrositos son incluso capaces de producir
algunos neurotransmisores. Además sabemos, por ejemplo, que ayudan a controlar la
cantidad de glutamato.
Con todos los datos comentados hasta el momento, ya se podrían enumerar las
funciones principales de los astrocitos:
• Constitución de la barrera hematoencefálica.
• Control del metabolismo neuronal.
• Producción de neuromoduladores y neurotransmisores.
• Metabolización de algunos neurotransmisores y neuromoduladores.
12
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El astrocito tiene gran capacidad de división, por ello, es capaz de reparar
cualquier lesión del sistema nervioso, formando una especie de “cicatrices”. La parte
buena, es que cierran el SN, pero presentan un invonceniente. Es posible que la cicatriz
de astrositos se retraiga con el tiempo y estimule las neuronas próximas, produciendo
focos epileptógenos. Además, dicha cicatriz, impide la regeneración de las fibras
nerviosas en algunas ocasiones (no pueden pasar a través de esa conjunción
astrocitaria).
Los astrositos van a ser elementos plásticos del sistema nervioso, de forma que
les da estructura (en caso contrario sería casi líquido). Las células satélites realizan la
misma función que los astrositos en el sistema nervioso periférico.
OLIGODENDROCITOS
Tienen una función primordial, que es producir mielina, que no es más que la
membrana de la célula del oligodendrocito que envuelve a las neuronas. Esa membrana
es un sistema aislante de las neuronas. De esta manera todas las células van envueltas en
una capa de fosfolípidos de mielina.
Clásicamente se han separado las fibras nerviosas según la cantidad de mielina que
existe a su alrededor, de forma que existen dos tipos principales:
a) Fibras que poseen mielina (mielínicas)
b) Fibras nerviosas que no tienen mielina (amielínicas).
No obstante cuando se observaron al microscopio electrónico, se observó que las
fibras amielínicas también poseían una capa aislante de fosfolípidos. Por ello, en la
actualidad, y aunque las fibras se siguen clasificando así, se incluyen en uno u otro
grupo en función del grosor, que es proporcional a la cantidad de mielina que poseen.
En las fibras mielíncas, la célula da vueltas al axón, mientras que en las amielínicas,
una sola célula rodeará a muchos axones. En el SNP estas células se conocen con el
nombre de células de Schwann, con un núcleo más o menos prominente y una
membrana que da vueltas abrazando al axón. Así, en el SNP, una misma neurona está
envuelta por muchas células de Schwann separadas por un espacio microscópico que se
conoce con el nombre de “nodo de Ranvier”. Ese espacio permite el intercambio iónico,
de ahí que la conducción en las fibras mielínicas sea “saltatoria”.
13
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VELOCIDADES DE CONDUCCIÓN Y CLASIFICACIÓN DE LAS FIBRAS
La velocidad de conducción de una fibra nerviosa es proporcional al grosor, de
forma que podemos emplear la medida de dicha velocidad para hacer una clasificación
de las fibras nerviosas. Cabe tener en cuenta, no obstante, que la fórmula que relaciona
velocidad de conducción y grosor de la fibra, es la que a continuación se muestra:
Normalmente, la velocidad en metros por segundo, vendrá determinada por una
constante a la que se le atribuye el valor “5”, valor que se verá afectado cuando existan
afecciones de fibras nerviosas (se reduce).
Atendiendo al grosor de las fibras nerviosas, se establece la clasificación de
Lloyd y Gasser. Cabe señalar sin embargo, que aunque ambas se emplean para fibras
sensitivas y motoras, la de Lloyd es más empleada para fibras sensitivas.
NO
TIPO DE VAINA MIELINIZADAS
MIELINIZADAS
Diámetro
22 µm 1,5 µm < 2 µm
de la fibra
Clasificación
I: Erlanger y 120 60 50 30 4 0,5
Gasser
Aα Aβ Aγ B C
Subclases:
Motoras para Fusomotoras Fusomotoras Autonómicas Autonómicas
músculo (a los husos) preganglionares postgangionares
Eferentes
esquelético Colaterales de
las fibras A
Aδ C
Aα y más pequeñas Cutáneas, Cutáneas,
Músculos y tendones: cutáneas musculares, musculares,
Aferentes
viscerales, etc. viscerales
I II III IV
a) Terminaciones Terminaciones secundarias Terminaciones Terminaciones
II. Lloyd primarias en en husos libres libres
Aferentes, husos Terminaciones
Musculoesqueléticas b) Terminaciones paciniformes
en tendones
14
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
De esta manera, Gasser realiza la división en fibras A, B y C, mientras que
Lloyd las divide en tipo I, II, III y IV. En la tabla que se presenta podemos observar
cómo la velocidad y el grosor son proporcionales, además del hecho de que todas las
fibras no mielinizadas tienen un grosor que nunca supera las dos micras.
CÉLULAS EPENDIMARIAS
Se trata de células que tapizan el interior de todas las cavidades del Sistema
Nervioso Central. Están formados por una sola capa de células de tipo cilíndrico con
cilios, que en ciertas localizaciones se especializan en la producción de hormonas,
además de quedar alguna célula madre en medio de las células. De esta manera, este
tipo de células sirven de barrera entre el líquido de las cavidades y el resto del SN, pero
es necesario aclarar también que estas células no son iguales en todo el conjunto del
Sistema Nervioso, pues presentan diferencias locales.
Debemos señalar, igualmente, que existen células ependimarias capaces de
segregar neuropéptidos en el LCR, por el que pueden difundir y actuar en lugares
lejanos de donde se han producido. Pero para que puedan actuar, estas sustancias deben
pasar del LCR al sistema nervioso, lo que tiene lugar en ciertas regiones, como es el
caso de la Región Postrema de intercambio de sustancias entre LCR y SN. Esta
capacidad de producir y transportar neuropéptidos se denomina neurocrina.
LA MICROGLÍA
Debido a su localización en el SN, se consideraba que eran células de estirpe
glial, de ahí el nombre clásico de microglía. Sin embargo hoy sabemos que se trata de
células de tipo defensivo y de origen mesodérmico.
LOS NERVIOS PERIFÉRICOS
Se trata de prolongaciones neuronales, fibras nerviosas al fin y al cabo. Pero las
neuronas, dependen de un sistema glial que les permitan mantenerse vivas, de ahí que
las fibras nerviosas posean también una glía propia. Esta glía está constituida por las
células de Schwann, que no son más que envueltas mielínicas. Finalmente, en el SNP
nos encontramos con una organización específica de tejido conjuntivo y, acompañando
15
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
al mismo, una serie de vasos (arterias, venas y capilares linfáticos). Esto supone otra
diferencia con el SNC, que no tiene sistema circulatorio linfático (su función la realiza
el LCR).
COMPONENTES DE LOS NERVIOS PERIFÉRICOS
Teniendo en cuenta que el sistema nervioso periféricos está compuesto por fibras
nerviosas, células de la glía y tejido conjuntivo, será preciso estudiar cada una de esas
partes por separado.
LA FIBRA NERVIOSA
La mayor parte de los autores afirman que los nervios periféricos son
únicamente axones, pero algunos de ellos sostienen la hipótesis de que en los nervios
sensitivos aparecen también dendritas especializadas que se comportan a modo de
axones.
Según los tipos de fibras nerviosas que contengan, existen tres tipos de nervios
periféricos:
1. Motores: existen tres tipos fundamentales de fibras nerviosas bien
diferenciadas:
a. Fibras motoras voluntarias: se encargan de los movimientos
de relación. Todas las fibras voluntarias se sitúan dentro del
SNC, en el tronco del encéfalo o agrupadas formando núcleos
motores. Los núcleos motores suelen estar situados en la
parte anterior del SNC, mientras que las partes
posteriores suelen ser sensitivas.
b. Fibras motoras involuntarias: forman parte del sistema
vegetativo, y se encargan de controlar las funciones de los
órganos vitales.
c. Fibras musculares cardíacas: tienen un sistema propio que
permite el latido, el sistema cardionector, que genera
estímulos para que se produzca la contracción.
16
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
2. Sensitivos: recogen información desde los órganos de los sentidos o
desde los órganos internos, y la dirigen al SNC para que sea
procesada.
3. Mixtos: se consideran mixtos aquellos nervios que están constituidos
tanto por fibras sensitivas como por fibras motoras.
VAINA DE MIELINA
No es más que la célula de Schwann que se envuelve sobre sí misma alrededor
de un axón. Existen nervios que presentan recubiertas mielínicas muy gruesas, mientras
que otros presentan una cantidad de mielina mucho menor.
TEJIDO CONJUNTIVO
Rodea al nervio, efectuando una acción de protección y sostén de las estructuras
nerviosas. Alrededor de cada fibra, va una capa de mielina, y finalmente, el tejido
conjuntivo.
En primer lugar, existe una capa de tejido conjuntivo que envuelve a cada uno de
los axones, y que recibe el nombre de Endoneuro. Posteriormente, se diferencia una
segunda envuelta que aísla un determinado número de fibras, formando fascículos, y
que denominamos Perineuro. Varios fascículos unidos se agrupan formando un nervio
rodeado por una capa de tejido conjuntivo que recibe el nombre de Epineuro.
El epineuro tiene características especiales, pues es un tejido conjuntivo muy
laxo que permite que los nervios, al moverse las
diferentes partes del cuerpo, se desplacen por las
estructuras próximas. Tiene especial importancia en
localizaciones donde el nervio pasa por zonas estrechas
o zonas relacionadas con huesos, como es el caso del
nervio cubital al pasar por el codo, o el mediano en el
canal carpiano.
17
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VASOS SANGUÍNEOS Y LINFÁTICOS
Los vasos, acompañan al tejido conjuntivo, de forma que nos encontraremos con
arterias y venas, así como con capilares linfáticos. Por ello, afecciones en el tejido
conjuntivo puede afectar a los vasos y a las fibras nerviosas.
NERVIOS SENSITIVOS
Los nervios sensitivos dependen siempre de una neurona, pero con un soma que
se localiza fuera del SNC, formando agrupaciones somáticas que se denominan
Ganglios Sensitivos. En el caso de los pares craneales, los ganglios suelen encontrarse
cerca de los orificios que permiten la entrada en el cráneo.
La médula espinal posee también una serie de raíces sensitivas propias y
dependientes de ella, que se agrupan en la parte posterior formando ganglios. Las
neuronas que forman estos ganglios se denominan neuronas pseudomonopolares, pues
de una única prolongación salen dos ramas: una que se dirige a la parte posterior de la
médula, y otra que se dirige a la perifería. De estas dos prolongaciones, una es un axón
y las otras unas dendritas. Pero las dendritas (que son las prolongaciones periféricas)
son “especiales” y se comportan como axones. El axón llega hasta la médula desde el
ganglio sensitivo, que está formado por miles de somas neuronales unidos a las células
satélites homólogas de los astrositos del SNC.
Estos nervios sensitivos (al igual que ocurre también con los motores), presentan
un origen real y un origen aparente. El origen real, en los sensitivos, se localiza donde
termina la rama de la neurona pseudomonopolar; mientras que el núcleo real motor se
sitúa en el lugar en el que se sitúan los somas neuronales dentro de la médula. Los
orígenes aparentes de ambos son los lugares desde donde los vemos salir del SN.
SISTEMA NERVIOSO VEGETATIVO
Este sistema requiere dos neuronas, generalmente una que se encuentra dentro
del sistema nervioso, y otra fuera del mismo. Las neuronas que se encuentran fuera del
sistema nervioso se organizan en ganglios vegetativos motores.
Existen dos tipos de motoneuronas: las que van directamente a unas fibras
musculares determinadas y que sirven para contraer todo el músculo (motoneuronas
18
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
alfa), y las que van a una estructuras específicas dentro del músculo que se conoce con
el nombre de husos neuromusculares (motoneuronas gamma).
La sensibilidad vegetativa depende de una neurona pseudomonopolar que recibe
la información de los distintos aparatos y la envía al SNC, mientras que la respuesta
motora requiere dos neuronas: una que parte desde la médula espinal y llega hasta el
ganglio, donde hace sinapsis con una nueva neurona motora que llegará directamente
hasta el órgano que tenga que realizar la respuesta correspondiente. Tanto las fibras
simpáticas como las parasimpáticos precisan de dos motoneuronas para responder a los
estímulos internos del organismo. No obstante, en el caso de los ganglios
parasimpáticos, éstos suelen localizarse próximos a los órganos a los que las
motoneuronas definitivas llevaran el impulso eléctrico.
LESIONES EN NEURONAS Y NERVIOS PERIFÉRICOS
Por norma general, el SNC no suele tener capacidad de regeneración, pero si
existe la posibilidad de que otras neuronas envíen sus prolongaciones al lugar donde ha
desaparecido una de ellas, intentando suplir de esta manera la deficiencia funcional que
supone su pérdida.
En cuanto al SNP, cabe decir que puede presentar diferentes patologías. Las
principales lesiones del SNP son las que a continuación se presentan:
• Neuroapraxia: es la pérdida de conducción de una fibra nerviosa. En estos casos
no hay alteraciones anatómicas. La única complicación es la pérdida de la
función del nervio. La lesión no deja secuelas, pues llega a curarse
completamente.
• Axonotmesis: en este caso existe lesión en los axones, la fibra está degenerada.
La gravedad dependerá proporcionalmente de la proximidad al soma neuronal.
Si la lesión no es próxima, se producirán dos fenómenos: en primer lugar surgirá
un proceso de degeneración (se mueren unos milímetros en la zona en
comunicación con el soma, así como todo el axón que no está en comunicación
con el susodicho soma). La mielina también degenerará. Después de este
fenómeno de degeneración, que dura unas semanas, comienza el período de
regeneración, que puede llevarse a cabo mediante dos procesos:
o Regeneración por yema terminal: en el extremo donde se produjo la lisis
aparece una yema terminal que crece por el tubo hasta que llega al
19
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
órgano diana correspondiente. En un principio es un axón muy delgado
sin mielina, que se engrosará y remielinizará. Esa yema terminal crece
entre 1-2 mm cada 24 horas, de ahí que sea necesario medir la distancia
de la lesión al punto donde termina el nervio para estimar el tiempo
necesario para que se complete la regeneración.
o Regeneración Colateral: si en las proximidades de la lesión existe un
nervio sano, éste enviará colaterales que se introducirán en los tubos que
ha dejado vacíos la lesión inicial.
• Neurotmesis: en esta lesión está afectado no sólo el axón, sino todas las capas
del nervio. El período de degeneración se sucede igual que en la axonotmesis.
Las posibilidades de regeneración también son:
o Regeneración por yema terminal: en estos casos, los “tubos” están
abiertos, de forma que existen tres posibilidades de actuación de esa
“yema terminal”:
Siga la dirección del tubo: dará al órgano diana y la lesión se
reparará.
Que se diriga por otro tubo de mielina y tejido conjuntivo que se
encuentre próximo: el nervio se dirige a lugares donde antes no
estaba. (Neurotización: aproximación de unos nervios periféricos
a otros para conseguir la recuperación de la movilidad de ciertos
músculos).
Formación de un ovillo por fuera de los tubos: cuando esto
sucede, puede morir la fibra nerviosa o recubrirse de tejido
conjuntivo y formar neuromas. Estos neuromas se descubrieron
por primera vez en personas con algún miembro amputado, pues
podían sentir los dedos que ya no tenían. Esto se debe a que las
fibras que se dirigían a estos lugares, formaban neuromas.
o Regeneración colateral: si en las proximidades de la lesión existe un
nervio sano, éste enviará colaterales que se introducirán en los tubos que
han dejado vacíos la lesión inicial.
• Axonoestenosis: es una compresión de un nervio periférico, bien de forma
aguda, bien de forma crónica. Esta patología se da muy a menudo, es la lesión
periférica más frecuente, y se da sobre todo en el nervio mediano en el túnel
carpiano o a través de los fascículos del pronador redondo, el cubital en el
20
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
conducto olecraniano, el nervio cutáneo poplíteo externo en el cuello del
peroné,…. Cuando los nervios son comprimidos, comienzan a tener dificultad
para que las sustancias que se forman en el cuerpo de las neuronas lleguen al
final de las prolongaciones, de ahí que el nervio empiece a engrosarse en las
zonas proximales y a adelgazar en las distales. Si este fenómeno actúa durante
mucho tiempo, llega un momento en que degenera el nervio completamente, y
se produce una lesión definitiva del mismo. Si se elimina la presión antes de la
degeneración del nervio, éste se recupera, de ahí la gran importancia del
diagnóstico precoz.
A causa de la existencia de toda esta patología, cobra especial importancia la
microcirugía de los nervios periféricos, que pretende aproximar los fragmentos
lesionados para producir la regeneración.
21
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La médula espinal es un órgano cilindroide, aplanado levemente de adelante
atrás. No obstante, no tiene una forma regular, sino que presenta partes más o menos
gruesas, que se denominan engrosamientos (uno lumbar y otro cervical) que están en
relación con el desarrollo nervioso de las extremidades. El engrosamiento lumbar, se
continúa con el cono medular. A éste le sigue el Fillum Terminale, ligamento de
sostén de la médula que terminará en el cóccix y la piel interglútea.
Para cuantificarla de alguna manera, podemos decir que presenta una longitud
que oscila entre los 40 y los 45 CM (se
extiende desde la articulación occípito-
atloidea hasta la articulación entre L1-L2),
mientras que el grosor ronda el centímetro.
En cuanto a su morfología, en la
médula se observa un surco que se conoce
con el nombre de Surco Medio Anterior.
Igualmente, existe otro en la parte posterior
de la misma que se denomina Surco Medio
Posterior. El surco medio posterior es
mucho menos marcado y se continúa con un
cordón de neuroglía, mientras que por el
surco medio anterior discurre la arteria
espinal anterior.
La médula presenta también el surco colateral posterior y el surco colateral
anterior, de los que salen los pares raquídeos. Las raíces raquídeas son los orígenes
aparentes de los nervios que salen de la médula espinal (pares raquídeos). Estos
nervios hacen un total de 31 pares, que clasificamos en:
• 8 Cervicales.
• 12 Dorsales o Torácicos.
• 5 Lumbares
• 5 Sacros
• 1 Coccígeo
22
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CAVIDADES
La médula espinal presenta una única cavidad alargada (conducto) en su parte
central interior, que recibe el nombre de Conducto del Epéndimo, y que se encuentra
tapizado por el epitelio ependimario.
SUSTANCIA GRIS Y SUSTANCIA BLANCA
En la médula espinal, la sustancia gris se sitúa en el centro, adquiriendo una
forma de “H”, con unas astas posteriores y unas anteriores. A estas astas anteriores y
posteriores hay que añadirles unas astas laterales que se presentan únicamente en la
región dorsal. En cuanto a funciones, cabe señalar que las astas posteriores son
sensitivas, mientras que las anteriores son las encargadas de la función motora.
En relación con la sustancia blanca, podemos comentar que se encuentra
rodeando la sustancia gris.
NOMENCLATURA DE LOS PARES RAQUÍDEOS
Los pares raquídeos suelen llevar el nombre de la vértebra inmediatamente
inferior, salvo en los dorsales, donde la terminología cambia, y el primer par dorsal es el
que sale por debajo de D1.
COLA DE CABALLO
Es el nombre que recibe el conjunto formado por el Fillum Terminale y los
pares raquídeos que van desde la articulación
entre L1-L2. Toda la superficie está recubierta por
líquido cefalorraquídeo. Esto tiene gran importancia
clínica, pues para extraer LCR (o introducir
medicación o contrastes radiológicos) se realiza una
punción lumbar para entrar en el saco dural,
siempre con una posición adecuada del paciente
23
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
(flexión de la columna), para conseguir abrir el paso entre las espinosas. Cuando
realizamos la punción atravesamos el ligamento supraespinoso, el interespinoso y el
ligamento amarillo. Acto seguido, entramos en contacto con la duramadre.
Es de importancia clínica recordar que, cuando llevemos a cabo una punción
lumbar, debemos asegurarnos de que no exista un aumento de la presión del LCR, lo
que se comprueba observando el fondo del globo ocular. Otra de las precauciones que
se deben tomar, es medir la presión del LCR con un manómetro a través de la aguja con
la que realizamos la punción. El problema que puede presentar el no realizar estas
comprobaciones es que, si existe un aumento de la presión hidrostática y ésta baja
mucho de forma repentina, es posible que se produzca una hernia de alguna parte del
encéfalo por el agujero occipital, llegando a provocar incluso la muerte del paciente.
LAS RAÍCES RAQUÍDEAS
Las raíces raquídeas son nervios periféricos, habitualmente mixtos, formados por
una parte sensitiva y otra motora. La parte motora de una raíz raquídea está formada por
motoneuronas que se encuentran en las astas anteriores. La parte sensitiva la conforman
neuronas pseudomonopolares que se encuentran en un ganglio sensitivo y que envían
fibras hacia las astas posteriores y hacia el nervio.
Los nervios raquídeos mixtos ya constituidos, van a salir del conducto raquídeo
por los agujeros de conjunción. Cuando salen del conducto, los pares raquídeos dan
cuatro ramas, de las cuales la más importante es la anterior. De este nervio primitivo
parten, igualmente, una rama más pequeña que es posterior; una tercera que es
recurrente (nervio sinuvertebral, o recurrente meníngeo), que vuelve a entrar e inerva
las paredes del SN por dentro; y una cuarta que constituye el nervio Rami-
comunicante blanco, que se dirige a unos ganglios vegetativos simpáticos situados
próximos a la médula. Estos nervios llevan fibras vegetativas al propio ganglio (son los
axones de las primeras neuronas de la vía vegetativa).
Las ramas posteriores de los pares raquídeos inervarán los músculos de los
canales vertebrales, también conocidos como los extensores del tronco o de la columna
vertebral. También poseen una rama que inervará la región dorsal del tronco. Estas
ramas, tienen más o menos importancia según el nervio, dado que tienen una patología
propia. En la chanela lumbosacra (de L1-S1) adquieren una mayor relevancia, puesto
24
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
que estas ramas tienen un importante componente sensitivo, lo que puede provocar
dolor lumbar.
Las ramas anteriores de los pares raquídeos, son aquellas que forman plexos
mediante anastomosis, y que van a inervar la musculatura ántero-lateral del tronco y los
miembros. A veces quedan aisladas segmentariamente (región dorsal), formando los
nervios intercostales.
ENGROSAMIENTOS
La cantidad de neuronas en la médula espinal es muy variable. Así, la sustancia
blanca aumenta a medida que ascendemos en la médula espinal. Esto se debe a que la
médula está en comunicación con el encéfalo y para comunicarse hacen falta fibras, que
partirán desde el propio encéfalo y que llegarán hasta la región sacra, dejando
prolongaciones nerviosas en cada nivel medular (de ahí que la cantidad de axones-
sustancia blanca- disminuya a medida que descendemos en la médula).
En los primeros momentos del desarrollo, la médula ocupa todo el conducto
raquídeo, pero a medida que pasa el tiempo, la columna vertebral crecerá más rápido
que la médula, de ahí que exista un “ascenso aparente” de la médula espinal. La
consecuencia es que, al principio, las raíces raquídeas salen de forma horizontal a través
de los agujeros de conjunción, pero al ascender la médula, se vuelven más verticales, y
forman la cola de caballo (siguen partiendo de los mismos agujeros de conjunción).
25
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
SEGMENTO MEDULAR O MIELÓMERO
En la médula espinal no existen surcos transversales, de ahí que nosotros, como
médicos, tengamos que buscar un sistema para segmentar la médula y poder establecer
un sistema de localización.
De esta manera, se establecen los mielómeros empleando las raíces nerviosas, de
forma que un segmento medular será aquella porción de la médula de donde sale cada
uno de los pares raquídeos. De esta manera tenemos 31 segmentos medulares, que se
dividen en:
• 8 segmentos cervicales
• 12 segmentos dorsales
• 5 segmentos lumbares
• 5 segmentos sacro
• 1 segmento coccígeo
Hay que señalar que los segmentos medulares no coinciden con las vértebras, de
ahí que sea necesario el empleo de una topografía vertebro-medular. Este problema se
soluciona con las existen Fórmulas de Chipault, que se basan en la localización de la
apófisis espinosa de una vértebra. Así, tenemos las siguientes directrices para localizar
los mielómeros (V= vértebra identificada a través de su apófisis espinosa):
• A nivel cervical: Sm= V+1
• A nivel dorsal superior ( de la primera a la sexta dorsal): Sm= V+2
• Dorsal inferior (desde la séptima a la onceava dorsal): Sm= V+3
• Vértebra D12: se corresponde con los segmentos medulares L3-L4-L5
• Vértebra L1: se corresponde con los mielómeros Sacros (1-5) y Coccígeo
SUSTANCIA BLANCA DE LA MÉDULA ESPINAL
La sustancia blanca de la médula espinal está dividida en cordones, de forma que
existen:
• Cordón anterior.
• Cordón posterior.
• Cordón lateral Derecho.
• Cordón Lateral Izquierdo.
26
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
A nivel cervical superior existe otro cordón a nivel posterior: el Cordón
Paramedio Posterior, que divide el cordón posterior en dos fascículos: el del Goll y el
de Burdach.
RECUBRIMIENTO MENÍNGEO DE LA MÉDULA
Según la división clásica de Bichat existen tres recubrimientos de los tejidos
nerviosos comunes a todo el SNC. Son las meninges, grupo entre las que se cuentan:
1. La Duramadre: es una cubierta de tejido conjuntivo con mucho colágeno que
tapiza todo el interior del conducto raquídeo para convertirlo en un continente
estanco. Así, se pega a las paredes del conducto, y se liga al tejido conjuntivo de
todo lo que entra o sale del sistema nervioso (nervios, arterias, venas,….). Entre
la duramadre y las paredes del conducto hay un espacio virtual que se conoce
con el nombre de espacio epidural, en el que nos encontramos dos cosas
prácticamente de forma permanente: un plexo venoso (plexos venosos
intrarraquídeos), muy importante dado que es el resultado de una anastomosis
funcional entre los sistemas de la cava superior y la inferior; y una gran cantidad
de grasa (tejido adiposo). A causa de la existencia de este espacio, es posible
que la duramadre se desprenda del conducto como resultado de patologías, como
es el caso de los hematomas epidurales. En ese espacio es donde se infiltra
también la anestesia epidural.
2. La Piamadre: es una capa mucho más fina y delicada, que tapiza todas las
estructuras que se encuentran dentro de la duramadre. Se introduce en todos los
surcos (médula, raíces, vasos,….), de forma que la piamadre es una lámina que
acompaña a cualquier elemento que entre o salga de la médula espinal. Como
todos los vasos que llegan están recubiertos de piamadre, decimos que la esta
capa es una lámina portadora de vasos. Cabe decir, sin embargo, que la
duramadre no está tapizada por la piamadre, sino por una envuelta propia que es
la aracnoides.
3. La Aracnoides: se trata de una envuelta que recubre la duramadre por su parte
interna y que presenta dos tipos de fibras: por un lado están las propias fibras
que tapizan la duramadre, y por otra, una serie tabiques que envía hacia la
médula (son pies fibrosos). A causa de la existencia de estos pies, a esta capa se
le atribuyó el nombre de aracnoides. Existe un tabique aracnoideo muy
27
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
importante que se denomina Ligamento en sierra o dentado, que se fija en la
piamadre y llega hasta los límites del conducto raquídeo, pero “salta” en los
agujeros de conjunción para dejar paso a los nervios (dando esa apariencia de
borde dentado).
ESPACIO LEPTOMENÍNGEO O SUBARACNOIDEO
Es el espacio que queda entre la aracnoides y la piamadre, y que está ocupado
por el LCR. De esta manera, se mantiene la idea de que el LCR ocupa tanto las
cavidades del SNC como la perifería del mismo.
VASCULARIZACIÓN DE LA MÉDULA ESPINAL
El tejido nervioso es dependiente de oxígeno y glucosa, por lo que requiere
aporte sanguíneo, que se realiza mediante arterias y venas. Es de interés señalar que el
SNC carece de vasos linfáticos.
Las arterias que llevan la sangre en la médula espinal son las Cuatro Arterias
Espinales, dos espinales anteriores, y dos espinales posteriores. Las dos arterias
espinales anteriores se juntan y se unen, y forman la Arteria Espinal Anterior. Además
de ellas, se pueden encontrar las arterias radiculares, de las que existe un gran número,
pues al comienzo del desarrollo existe una arteria radicular por cada raíz nerviosa, pero
28
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
su número va disminuyendo a medida que se avanza en el desarrollo. Existen dos
arterias radiculares especialmente importantes, que son la Arteria de Engrosamiento
Cervical y la Arteria de Engrosamiento Lumbar (arteria de Adankiewich).
Todas las arterias del sistema nervioso central (por norma general), se anastomosan
formando una red, que está situada a nivel de la piamadre. No obstante, a partir de la red
anastomótica de superficie que se forma en la perifería de la médula, parten las Arterias
Intramedulares, que al igual que todas las arterias que penetran en el sistema nervioso
central, se trata de ramas terminales, por lo que un fallo en ellas provoca la muerte de
regiones específicas del SNC (sólo hay pequeñas anastomosis no funcionales). Estas
arterias intramedulares, pueden dividirse en dos grandes grupos:
• Arterias Centromedulares: son un grupo de arterias que dependientes de
la arteria espinal anterior que se dirigen a la zona central de la médula
espinal.
• Arterias radiales: se trata de arterias que parten de la red anastomótica y
se dirigen a la perifería de la médula espinal.
La repartición de la vascularización es importante para la práctica clínica, pues
en ocasiones puede aparecer necrosis en distintas localizaciones medulares en función
de la región arterial afectada. Por ejemplo, si existen problemas en las arteria espinal
29
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
anterior, se producirá necrosis de la zona central de la médula, dando como resultado
una cavidad muy grande y una clínica especial.
No existe un número de vasos igual en toda la médula pues hay zonas de mayor
y menor vascularización. La zona de menor vascularización se sitúa en la región dorsal,
y cuando hay un déficit vascular puede haber lesiones de la médula simplemente por
esta carencia.
De esta manera sabemos que no es precisa una luxación de columna para
provocar una lesión, basta con una flexión repentina del tronco que provoque un
estiramiento rápido de la médula o con la obstrucción de un vaso a causa de algún
traumatismo.
Las venas de la médula se dirigen a la perifería formando las venas radiadas y
centromedulares, y van a acabar en unos grandes plexos venosos situados dentro del
raquis que son los plexos venosos intrarraquídeos, que acabarán por desembocar en la
vena cava superior o en la cava inferior.
30
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En esta unidad temática nos vamos a central en el estudio de una estructura
nerviosa que se encuentra entre la base del cerebro y la médula espinal, y que se conoce
con el nombre de Tronco del encéfalo. Las partes de su estudio en las que nos vamos a
centrar son las que presentamos a continuación.
MORFOLOGÍA EXTERNA DEL TRONCO DEL ENCÉFALO
A) Anatomía Microscópica y morfología general del Tronco encéfalo (TE):
a. Concepto
b. Partes
c. Límites
d. Generalidades de la configuración interna del TE.
B) Conformación Exterior.
a. Morfología de la cara anterior del:
i. Bulbo
ii. Protuberancia
iii. Mesencéfalo
b. Morfología de las caras laterales de:
i. Bulbo
ii. Protuberancia
iii. Mesencéfalo
c. Morfología de la cara posterior del TE.
C) Morfología Externa del cuarto ventrículo
a. Límites del cuarto ventrículo.
b. Suelo y techo del cuarto ventrículo.
31
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
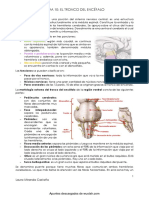
CONCEPTO DE TRONCO ENCEFÁLICO
Es el segmento inferior del encéfalo que se sitúa delante del cerebelo y por
debajo de la base del cerebro. Constituye el punto de unión entre médula y encéfalo,
está en relación con el occipital y se va a situar delante de la fosa cerebelosa.
PARTES DEL TRONCO
Consta de tres pisos superpuestos en sentido cráneo-caudal, de tal manera que el
primero de ellos sería el bulbo, que posee gran importancia funcional debido al enorme
número de centros vitales que en él existen, y que tiene un parte de cilindro inferior que
se une con la médula con una zona más aplanada en la parte superior. El segundo piso o
intermedio lo constituiría el punte o protuberancia, que se puede comparar con la forma
de un cubo, y que tiene una cara anterior, una posterior y dos laterales. Finalmente, el
último piso lo conforma el mesencéfalo, y estaría constituido por dos estructuras: los
pedúnculos cerebrales a nivel anterior, y los tubérculos cuadrigéminos en la posterior.
El límite inferior del tronco del encéfalo es el lugar con el que entra en contacto
con la médula y esto tiene lugar en la Decusación de las pirámides o de Mistichelli. El
límite superior lo formarían el sistema de Vías Ópticas, que está formado en primer
lugar por el quiasma (punto donde se cruzan las fibras del nervio óptico) y las cintillas
ópticas, que son la continuación del quiasma y que van a bordear por ambos lados al
TE.
Otras estructuras que nos sirven para separar las partes del TE son el surco
bulbo-póntico, que separa bulbo y puente, y el surco ponto-mesencefálico, que como su
nombre indica, separa el puente del mesencéfalo.
En la cara posterior del TE, observaremos que el bulbo y la protuberancia o
puente se abren al cerebelo, algo en lo que no se incluye el mesencéfalo. Además, el
bulbo y la protuberancia van a formar el suelo del cuarto ventrículo.
32
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
GENERALIDADES DE LA MORFOLOGÍA INTERNA DEL TRONCO DEL
ENCÉFALO
En el TE aparecen núcleos o acúmulos de núcleos que van a controlar la médula,
así como la estructura interna del TE. De todas formas se puede establecer un
paralelismo entre la organización de la médula espinal y del TE, de tal manera que esta
sustancia gris (los núcleos) se van a situar en el interior a nivel del TE, y la sustancia
blanca se va a situar en la perifería. Del TE van a emerger una serie de nervios que
pueden ser motores (si tienen origen en una neurona motora), sensitivos o mixtos. Estos
nervios son los doce pares craneales, de tal manera que se nombran con el número más
bajo a los que emergen de la parte más superior del encéfalo. Estos pares craneales no
son periféricos, proceden y forman parte del SNC, y tendrán dos orígenes: uno real y
otro aparente. El origen real de los pares craneales sería, por una parte, el soma o el
cuerpo de la neurona que se localiza en el TE (y que en este caso es una neurona
motora) y que dará lugar a un nervio motor. Si es una neurona sensitiva, el origen real
sería el punto de unión del TE con la neurona sensitiva (recibe información). El origen
aparente sería el punto en la superficie del TE donde saldría o bien el nervio motor o
donde entraría el nervio sensitivo
CARA ANTERIOR DEL TRONCO ENCEFÁLICO
CARA ANTERIOR DEL BULBO
El bulbo, en su conjunto, tendría una forma de cono truncado, con el vértice
inferior uniéndose al cilindro que configura la médula, y con la base superior que se
uniría al puente o protuberancia. Va a presentar una cara anterior, una posterior, y dos
laterales. Su límite inferior es la Decusación Piramidal. A lo largo de toda su cara
ventral, asciende un surco, continuación de otro surco de la médula, y que se conoce con
el nombre de Surco Medio Anterior. Este surco va a acabar en un agujero u orificio que
se denomina Agujero Ciego, que no es más que la unión entre el Surco Medio Anterior
y el surco bulbopóntico. A ambos lados del agujero ciego, tiene el origen aparente un
par craneal que es el Motor Ocular Externo (VI par craneal). El Motor Ocular Externo
33
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
es el encargado de la inervación extrínseca del globo ocular, pues inerva el recto
externo. A ambos lados del Surco Medio Anterior aparecen otros surcos que también
son continuación de los surcos de la médula, y que son los surcos colaterales anteriores,
que ascienden en sentido
vertical y son paralelos al
Surco Medio Anterior, y en la
cara más superior de la parte
anterior del bulbo, se desplazan
ligeramente hacia delante, para
dar lugar al surco preolivar,
que recibe este nombre porque
deja paso a una estructura
saliente a nivel del bulbo que
se denomina oliva bulbar. En
este surco preolivar, tendrá el origen aparente un nuevo par craneal que es el Hipogloso
(XII par), de ahí que se denomine también Surco del Hipogloso.
Finalmente, rematamos la cara anterior, diciendo que entre el Surco Medio
Anterior y los surcos preolivares, existen unas prominencias que se llaman pirámides
anteriores del bulbo, que son fibras que proceden de la corteza cerebral, y que
descienden también por la protuberancia o puente, haciendo prominencia en el bulbo, y
que parece que es la continuación de los cordones anteriores de la médula (aunque
realmente se trata de fibras motoras voluntarias). Estas pirámides anteriores del bulbo,
van a enmarcar más el surco del hipogloso.
CARA ANTERIOR DE PROTUBERANCIA O PUENTE DE VAROLIO
El límite inferior de la protuberancia se sitúa en el Surco bulbo-póntico, y el
límite superior en el surco ponto-mesencefálico.
La protuberancia va a tener una cara anterior, una posterior, y unas caras
laterales. Está formada por fibras transversales, que se va a continuar lateralmente con
las fibras del pedúnculo cerebeloso medio, que une el TE al cerebelo.
Si se realiza un corte transversal, observaríamos por una parte las fibras, y por
debajo, prodríamos ver los haces de la vía piramidal. Al unir las fibras estos dos haces,
34
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
se forma una hendidura o canal en el centro, que se denomina Canal Basilar (hendidura
que queda entre las fibras al pasar por debajo las vías piramidales). Por ese canal
basilar, va a discurrir una estructura que se denomina tronco basilar. El tronco basilar se
forma en el agujero ciego por la unión de unas arterias, las arterias vertebrales, y en la
porción superior, va a dar lugar a otras arterias, que serán las arterias cerebrales
posteriores. Las prominencias de las fibras que quedan a ambos lados del canal basilar,
se denominan protuberancias o rodetes piramidales (o protuberenciales).
No existe un límite claro entre la cara anterior y la lateral de la protuberancia.
Dicho límite lo marcará la salida aparente de un par craneal: el Trigémino (V par). El
trigémino tiene una raíz motora y una raíz sensitiva que están separadas por una porción
de sustancia blanca que se denomina Lígula de Wrisberg. Este par craneal va a dar
inervación a los músculos masticadores y va a ser el nervio sensitivo del globo ocular.
CARA ANTERIOR DEL MESENCÉFALO
La cara anterior del mesencéfalo va a estar limitada en su parte superior por el
Sistema de Vías Ópticas, y a nivel inferior, lo estará por el surco ponto-mesencefálico.
El mesencéfalo está constituido por los pedúnculos cerebrales y el espacio que éstos
35
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
dejan en medio, que se conoce como Espacio Interpeduncular. Los pedúnculos
cerebrales, en su ascenso al cerebro, dejan en medio un espacio triangular que es el
Espacio Perforado Posterior. La base de dicho triángulo, es la Fosa Interpeduncular. A
ambos lados de la línea media de este triángulo, se observa la salida aparente el III par
craneal, que es el Motor Ocular Común. El espacio perforado posterior, está constituido
por un montón de orificios que dan entrada a las ramas de la arteria cerebral posterior.
A un nivel más superior de este espacio, ya en el diencéfalo, observamos unas
prominencias en forma de tubérculos que se denominan Tubérculos Mamilares (uno
derecho y uno izquierdo). Por encima de dichos tubérculos mamilares, se diferencia una
prominencia en forma de embudo que se continúa con un tallo, y este tallo con una
glándula, que es la hipófisis. El embudo se corresponde con el infundíbulo, el tallo con
el tallo de la hipófisis, y la glándula, como es lógico, sería la hipófisis. Esta estructura
presenta una porción más saliente que es la Eminencia, que se sitúa en la parte más
posterior del infundíbulo.
CARAS LATERALES DEL TRONCO DEL ENCÉFALO
CARAS LATERALES DEL BULBO
36
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La cara lateral del bulbo es la continuación de la cara lateral de la médula, y los
límites de ésta serían el Surco Colateral Anterior y el Surco Colateral Posterior (que
aparecen también en la médula), de forma que dicha cara lateral podría considerarse la
continuación del cordón lateral de la médula. En medio de este cordón se va a formar
otro surco que es el Surco de los Nervios Mixtos, pues en él se distingue la salida
aparente de los siguientes pares craneales (nombrados en sentido cráneo-caudal):
• IX Par o Nervio Glosofaríngeo: da inervación a la musculatura faríngea.
• X Par o Nervio Vago: aporta inervación visceral de tronco (tórax) y cuello.
• XI Par o Nervio Espinal: tiene dos porciones, que son, por un lado, una salida de
raíces a nivel bulbar (encargada de la inervación de la musculatura de la laringe,
excepto el Crico Aritenoideo Posterior), y por otro una porción medular (que
inervará el esternocleidomastoideo y el trapecio).
En la parte superior, el surco de los nervios mixtos va a dar lugar al surco
retroolivar, que se sitúa en la parte posterior de la oliva bulbar. En este surco, podemos
encontrar la Arteria Retroolivar (o postolivar).
En la parte superior de la oliva bulbar, nos vamos a encontrar una depresión que se
conoce con el nombre de Fosita Supraolivar, lugar en el que tendrá lugar la salida
aparente del Nervio Facial (VII
par), así como del intermediario
de Wrisberg. El nervio facial, es
el encargado de la mímica de la
cara, y su parte sensitiva (el
intermediario) se hace cargo de
la inervación de las glándulas de
la boca y de la función gustativa
(salvo el sabor amargo, que
corre a cargo del glosofaríngeo).
Detrás de la fosita supraolivar, nos encontramos con la Fosita Lateral del Bulbo, de
donde saldrían las raíces del VIII par craneal, que presenta una función auditiva
realizada por la rama coclear y una función de equilibrio realizada por la rama
vestibular. El VII y el VIII par, se unen para entrar por el conducto auditivo interno y
forman el Paquete Acústico-Facial.
CARA LATERAL DE LA PROTUBERANCIA O PUENTE DE VAROLIO
37
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Sólo concretar que la cara lateral del puente o protuberancia se inicia a partir de
la salida aparente del quinto par, y que en ella, las fibras transversales de la
protuberancia se continúan con el pedúnculo cerebeloso medio, que servirá de medio de
unión entre la propia protuberancia y el cerebelo.
CARA LATERAL DEL MESENCÉFALO
Los pedúnculos cerebelosos unen las distintas porciones del tronco del encéfalo
con el cerebelo, forma que el superior establece la unión entre cerebelo y mesencéfalo,
el medio entre cerebelo y protuberancia y el inferior entre cerebelo y bulbo raquídeo.
En la cara lateral del mesencéfalo observamos cómo ascienden los pedúnculos
cerebrales. En dicho ascenso, los pedúnculos dejan un surco lateral y oblicuo que
asciende hacia arriba y que se conoce con el nombre de Surco Lateral del Istmo, que se
marca mucho a causa de la forma en la que sobresale la protuberancia. De esta
estructura diverge el Pedúnculo Cerebeloso Superior.
38
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Al mismo tiempo observamos que en la parte posterior del mesencéfalo tenemos
unas estructuras abultadas que se conocen con el nombre de Tubérculos Cuadrigéminos,
de los cuales, el superior, se va a unir a una estructura talámica (que es el Cuerpo
Geniculado Interno) a través del Brazo Conjuntivario Inferior. Estas tres estructuras dan
lugar al Triángulo de Reil.
Al otro lado del istmo se encuentra el Pedúnculo Cerebeloso Superior. Estas dos
estructuras divergen entre ellos formando una estructura triangular, que se conoce con el
nombre de Triángulo de Reil. Es muy importante, pues de él van a emerger unas fibras
de la sensibilidad auditiva, que se conocen con el nombre de Cintilla de Reil (emergen
en el Surco Lateral del istmo e irradian en el triángulo de Reil).
El triángulo de Reil está compuesto por una base anterior, que es el surco lateral
de istmo, un lado inferior, que sería la raíz externa del pedúnculo cerebeloso superior y
un tercer lado superior que sería el brazo conjuntival inferior. El tubérculo
cuadrigémino inferior se considera el vértice de dicho triángulo.
CARA POSTERIOR DEL TRONCO-ENCÉFALO
Para una correcta comprensión de la cara posterior del tronco encefálico,
estudiaremos en primer lugar las caras no ventriculares y posteriormente las caras que
forman parte del cuarto ventrículo.
CARAS NO VENTRICULARES
CARA POSTERIOR DEL BULBO
En un primer lugar nos encontramos con un Surco Medio Posterior que supone
la continuación del surco medio posterior de la médula. A ambos lados de dicho surco
podemos distinguir los surcos laterales posteriores, también continuación de la médula,
En el bulbo, aparece también el surco paramedio o intermedio. Del mismo modo, nos
encontramos unos haces que nos separa este surco paramedio: por un lado el haz del
Goll, y por el otro el Haz de Burdach. Existen también dos elevaciones a cada lado del
surco medio posterior: la interna se denomina clava o maza y la externa se conoce con
el nombre de Tuberculus Cuneatus.
39
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Los pedúnculos cerebelosos inferiores o cuerpos restiformes se separan y
delimitan a nivel inferior el cuarto ventrículo. Estos pedúnculos entran en comunicación
en un punto que se conoce con el nombre de Óbex o Cerrojo.
CARA POSTERIOR DEL MESENCÉFALO
Nos encontramos los pedúnculos cerebelosos superiores, que limitan la parte
superior del suelo del cuarto ventrículo; así como con una lámina y unas prominencias,
que son los tubérculos cuadrigéminos: dos de ellos anteriores o superiores y dos
posteriores o inferiores. Esta lámina de tubérculos cuadrigéminos está cruzada por un
surco que se conoce con el nombre de Surco Cruciforme. En la parte más alta de la rama
vertical de surco cruciforme nos encontramos con la Glándula pineal, y en la parte más
inferior de dicha rama nos observamos otra estructura que se denomina Frenillo de la
Válvula de Viussens. A ambos lados del frenillo tiene la salida aparente el IV par
craneal.
CARAS VENTRICULARES
EL CUARTO VENTRÍCULO
El cuarto ventrículo será dividido, para su correcto estudio, en suelo y techo del
mismo, y analizaremos por separado cada una de esas partes.
SUELO DEL CUARTO VENTRÍCULO
El cuarto ventrículo es
una estructura que estaría
formada en su suelo por parte
de la cara posterior del bulbo y
por la cara posterior del puente
y mesencéfalo. Este ventrículo
forma una especie de cuña
introduciéndose en el cerebelo,
40
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
de forma que se sitúa delante de éste.
El Suelo del Cuarto ventrículo tendría forma de rombo, en el que nos
encontramos un surco en sentido cráneo-caudal, que se parece al tallo de una pluma y
que se le da el nombre de Tallo del Calamus Scriptorius. Éste se dilata en su parte más
caudal, dando lugar a una estructura que se denomina Pico del Calamus.
Se pueden observar, igualmente, unas fibras que parten de los ángulos externos
del suelo de este cuarto ventrículo, y que reciben el nombre de Fibras Arciformes o
Estrías Acústicas. De estas fibras, hay una que se prolonga más y que se denomina
Barilla Armónica de Bergman.
Para comprenderlo mejor, podemos dividir el cuarto ventrículo en dos
triángulos: uno inferior o bulbar y uno superior o póntico.
En el triángulo inferior o bulbar aparece, en la parte interna, una estructura que
sobresale que es de color blanquecino que se conoce con el Ala Blanca Interna. Aquí
tendrá el origen real el XII par craneal y por ello se denomina Trígono del Hipogloso.
En la parte externa, nos encontramos otra porción saliente blanquecina de nuevo, que es
el Ala Blanca Externa. En ella existe un tubérculo donde hay núcleos vestibulares del
VIII par, de ahí que se denomine también Trígono Vestibular. Finalmente, nos queda
un triángulo de vértice superior y base ínfero-externa, que es más profundo que las
estructuras limitantes y tiene una coloración grisácea, de ahí su nombre de Ala Gris. En
esta ala gris, tiene el origen real el X par, de ahí que se conozca también con el nombre
de Trígono del Vago.
En el triángulo superior o póntico vamos a encontrarnos, a ambos lados del
Surco Medio, unas prominencias que se llaman Eminencias Teres. Si realizamos un
corte a nivel de las eminencias teres, observamos que son el resultado del rodeo que
hace el axón del facial al núcleo del Motor Ocular Externo. Continuando en vertical a
las Eminencias Teres, tenemos otras estructuras que son los Funículum Teres. En medio
de ambas eminencias, a nivel del
surco medio, tenemos la fóvea
media, donde iría el IV par. Se
distingue también un tubérculo que
queda por encima de la base del
Ala blanca Externa y que se llama
Tubérculo Acústico. A nivel lateral
41
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
superior, nos encontramos con la Fóvea superior, en la que se observa, por
transparencia, el V par. Al lado de la fóvea superior aparece un acúmulo de neuronas
negruzcas que se denomina Locus Coerelus (neuronas noradrenérgicas).
TECHO DEL IV VENTRÍCULO
El techo del cuarto ventrículo estaría formado por una vertiente superior y una
vertiente inferior. Estas dos vertientes, se unen en el cerebelo en un punto que se
denomina Núcleo Fastigium.
La vertiente superior estaría limitada por los pedúnculos cerebelosos superiores,
que se unen por el velo medular anterior o válvula de Viussens. La vertiente superior es
más gruesa que la vertiente inferior, sobre todo en la zona de la válvula de Viussens.
La vertiente inferior tiene una porción más gruesa relacionada con el cerebelo y
una más fina en contacto con el bulbo. Está limitada por los Pedúnculos cerebelosos
inferiores, y en él se pueden
observar dos estructuras
nerviosas en forma de semiluna
que son las Válvulas de Tarín,
que se sitúan a nivel lateral, y
que están pegadas al techo del
cerebelo. Existe también otro
orificio a nivel medio de la
vertiente inferior que es el
orificio de Magendi, que
sobrepasa la membrana tectoria y los plexos coroideos. Finalmente, tenemos otros dos
orificios a ambos lados de la vertiente inferior que comunican con el exterior, que son
los agujeros de Lushka, por los que salen, en muchos casos, los plexos coroideos, dando
lugar al Cuerno de la Abundancia o Canastilla de Flores (comunican el ventrículo con el
espacio subaracnoideo).
Cabe señalar, igualmente, que todo el techo del cuarto ventrículo está recubierto
por epitelio ependimario, que recubre todas las cavidades. De esta manera, en la
vertiente inferior podemos distinguir las siguientes capas que la recubren de dentro a
fuera:
42
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
a) La membrana tectoria: se trata de una membrana epitelial que recubre la parte
interna de los cuerpos restiformes o pedúnculos cerebelosos inferiores. Dentro
de la membrana tectoria nos encontramos unas estructuras nerviosas
rudimentarias que recubren o forman el Óbex o cerrojo, así como otra
estructura laminar que es la Lígula (que va a recubrir la cara interna de los
pedúnculos cerebelosos inferiores lateralmente y a ambos lados, e incluso
levemente de forma transversal)
b) Tela coroidea o formaciones coroideas: está compuesta por los vasos coroideos
tapizados con piamadre. Existen plexos longitudinales que recubren las partes
laterales de la vertiente inferior, y transversales, que son los que hacen los
recesos y dan lugar al cuerno de la abundancia.
43
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En el cerebelo y cerebro es un órgano que se encuentra situado en la fosa craneal
posterior o cerebelosa, y en la parte posterior del tronco del encéfalo, unido a éste
mediante los tres pares de pedúnculos cerebelosos. Su organización histológica muestra
la sustancia gris en la perifería, y la sustancia blanca en el centro. Una característica
especial que comparten cerebro y cerebelo, es que poseen gran cantidad de sustancia
gris en su corteza, por lo que son necesarios una serie de plegamientos de la misma. El
resultado, en el caso del cerebelo, es una distribución arboriforme de la sustancia blanca
en su interior, lo que los clásicos dieron en llamar “Árbol de la vida” (el cerebelo era
considerado como un órgano vital, afirmación totalmente incierta)
Desde el punto de vista de la anatomía macroscópica, podemos dar una serie de
datos morfométricos que nos ayuden a comprender la estructura del órgano. Así,
podemos decir que presenta unas dimensiones aproximadas de 5x5x10 (medidas en
sentido vertical, anteroposterior y horizontal). Observamos, por tanto, que tiene una
anchura que lo hace sobresalir lateralmente del tronco encefálico si observamos las
estructuras desde un punto de vista anterior. En cuanto a su peso, podemos dar una
medida aproximada de 140 gramos, pero con una variabilidad que llega hasta el 10%.
Desde el punto de vista funcional, debemos señalar que el cerebelo consta de
tres partes funcionales diferenciadas, y que a continuación enumeramos:
1. Arquicerebelo o cerebelo vestibular: relacionado funcionalmente con el
sistema vestibular (coordinación del equilibrio). Este tipo de cerebelo
aparece ya en animales acuáticos, donde los movimientos del tronco son
suficientes para mantenerse a flote.
2. Paleocerebelo: relacionado funcionalmente con el tronco encefálico y la
médula espinal. Se encarga de la coordinación de la postura y es propio de
animales terrestres.
3. Neocerebelo: relacionado funcionalmente con la corteza cerebral
(coordinación de los movimientos voluntarios).
Vemos, por tanto, que el cerebelo es un órgano de coordinación del movimiento
y es el elemento más importante para el aprendizaje motriz.
Debemos especificar que en cerebelo existen unas estructuras medianamente
centrales que se conocen con el nombre de núcleos cerebelosos, y que son cuatro:
44
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
1. El núcleo del Techo (arquicerebeloso)
2. El núcleo Globoso o Globuloso (Paleocerebeloso)
3. El núcleo Emboliforme (Paleocerebeloso)
4. El núcleo Dentado u Oliva del Cerebelo (Neocerebeloso)
El límite entre cerebro y cerebelo viene determinado por una lámina de
duramadre que los separa, y que recibe el nombre de Tienda del Cerebelo.
Observando macroscópicamente el cerebelo desde una vista inferior, se ve una
parte central que se denomina vermis, que está separando los dos hemisferios del
cerebelo. El cerebelo abraza el tronco del encéfalo gracias a una estructura que se llama
incisura semilunar, que se corresponde en la parte posterior con otro surco que se
conoce con el nombre de incisura marsupial.
Gracias a una fisura transversal en forma de semilunar que presenta el cerebelo,
podemos dividir al mismo en tres caras: una anterior, una superior y otra inferior (estas
dos últimas separadas por el surco horizontal).
Unas estructuras que es necesario señalar son las laminillas cerebelosas, fisuras
que recorren toda la corteza del órgano, así como las amígdalas cerebelosas, que se
observan en una vista posterior y por encima de las cuales se localiza el cuarto
ventrículo. Esta estructura tiene especial importancia, pues posee una patología propia,
que se denomina enclavamiento de las amígdalas, que es el resultado de una hernia del
Sistema nervioso resultado de un aumento de presión a nivel superior. Esto puede ser
muy grave, pues la amígdala puede ejercer presión sobre los centros vitales del bulbo
raquídeo y llegar a provocar la muerte del paciente.
OBSERVACIÓN DE LAS CARAS DEL CEREBELO
Para poder observar todas las caras del cerebelo es necesario realizar su
extracción del tronco del encéfalo, para lo que es preciso cortar los pedúnculos
cerebelosos.
Con estos cortes, en la cara anterior no encontraremos con los tres pedúnculos y,
uniendo a los superiores está la válvula de Viussens. Por encima de ella, queda una
porción de cerebelo que se conoce con el nombre de Língula. A nivel de los pedúnculos
cerebelosos inferiores nos encontramos con la Lígula. Limitante con la vertiente inferior
del techo del cuarto ventrículo existe una parte de cerebelo que recibe el nombre de
Nódulo.
45
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Recubriendo a todo el ventrículo existe un epitelio ependimario perforado por
una serie de orificios que son el de Magendie y los de Luschka, que ponen en contacto
la cavidad del cuarto ventrículo con el espacio subaracnoideo. Si se produce la oclusión
de estos orificios, el resultado será una patología conocida como hidrocefalia (aumento
de tamaño del sistema nervioso).
El cerebelo es convexo en todas sus direcciones y en su parte central se observa
el vermis (superior e inferior) del cerebelo. A ambos lados se encuentran los hemisferios
cerebelosos. La diferencia en el vermis superior e inferior reside en que el superior es la
zona más alta del cerebelo, y en la parte inferior, es la que menos resalta, pues están
muy abultados los hemisferios.
En el cerebelo pueden observarse también unos surcos transversales que lo
atraviesan, entre los que destacan algunos más gruesos que se denominan Cisuras (son
más constantes y profundos que los surcos comunes). Las cisuras, por norma general,
separan el cerebelo en lóbulos. Para poder observar esto con corrección, debemos
realizar un corte del cerebelo a nivel del vermis. Con este corte, y tal como se puede
observar en el esquema observamos distintos surcos y porciones cerebelosas:
• Cisura Prima o transversal superior: separa la parte más alta del cerebelo
(culmen) de la porción en la que empieza a descender (déclive).
• Cisura Póstero-Lateral: es aquella que separa el nódulo y el flóculo del resto del
cerebelo. Es tan profunda, que los antiguos consideraban que estas estructuras
46
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
no pertenecían al cerebelo. Hoy se sabe que son uno de los componentes más
importantes del arquicerebelo.
• Surco Prepiramidal: separa la pirámide del túber del cerebelo. Se denomina
prepiramidal porque en los primeros estadíos de desarrollo del cerebelo, se
encuentra delante de la pirámide, aunque una vez que crece, el surco quede por
detrás de la misma.
• Surco circunferencial u horizontal de Viq-D’Azir: nos separa la cara superior de
la cara inferior del cerebelo.
Los demás surcos que podemos nombrar, aunque su importancia sea mucho
menor, son los siguientes:
• Surco Precentral: separa la língula del lobulillo central.
• Surco Postcentral: separa el lobulillo central del culmen.
• Surco Semilunar o superior: separa el déclive del Folium
• Surco Postpiramidal: separa la pirámide de la úvula o campanilla.
En un principio, en el cerebelo podemos encontrarnos con cuatro lóbulos, que
englobarán distintas partes del cerebelo. Así, empleando una clasificación embriológica,
podemos agrupar las distintas partes del cerebelo de la siguiente forma :
a) Lóbulo anterior: engloba la Língula, el Lobulillo Central y el Culmen.
b) Lóbulo flóculo-nodular: formado por el Nódulo y el Flóculo.
c) Lóbulo medio: constituido por el Déclive, el Folium y el Túber.
d) Lóbulo posterior: que engloba la Úvula (o campanilla) y la Pirámide.
47
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
A nivel de los hemisferios, van a continuarse con los surcos del vermis, de forma
que la língula va a estar continuada por dos partes de cerebelo a los lados que se
denominan frenillos de la língula. A los lados del lobulillo central están las alas del
lóbulo central. A cada lado del culmen, en los hemisferios, aparecen unos lobulillos
que reciben el nombre de nódulos cuadriláteros anteriores. A nivel del déclive se
sitúan los lobulillos cuadriláteros posteriores. A nivel del folium localizamos los
lobulillos semilunares superiores, y finalmente, junto al túber, se encuentran los
lobulillos semilunares inferiores.
A la altura de la pirámide, en lugar de existir un solo lobulillo, existen dos a cada
lado: el digástrico y el gracilis o delgado. Esto se debe a que en los hemisferios existe
un surco más, que es el que se conoce con el nombre de surco semilunar inferior, que
hace que a ambos lados de la pirámide aparezcan dos lobulillos. A los lados de la úvula
están las amígdalas y acompañando al nódulo el flóculo.
FORMACIONES DE LOS TRES CEREBELOS
Una vez conocidas las distintas partes que componen el cerebelo, procederemos
a clasificarlas de acuerdo con las funciones que desempeñan:
a) Arquicerebelo: está compuesto por el nódulo y el flóculo. Algunos autores
introducen la língula, aunque la mayor parte de los estudiosos consideran que
esto es falso.
b) Paleocerebelo: compuesto por língula, lóbulo central, culmen, lóbulo
cuadrilátero, ala del lóbulo central pirámide y úvula.
c) Neocerebelo: el resto de las partes del cerebelo que aún no han sido nombradas
pertenecen al neocerebelo (casi toda la superficie de los hemisferios).
ÁNGULO PONTOCEREBELOSO
Es importante para la práctica médica, y se trata de un ángulo diedro que se
forma entre la cara lateral del tronco del encéfalo y la cara anterior del cerebelo. Es una
zona típica de tumores con una sintomatología muy típica. Los tumores empiecen por
vértigos y mareos (por opresión del VIII par), que siguen con problemas faciales (VII
par craneal) y posteriormente puede afectar también al trigémino, glosofaríngeo o
pneumogástrico (menos común).
48
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El cerebro es un órgano que se encuentra situado dentro del cráneo, protegido
por él. Podemos dividir el cerebro en dos partes principales: por un lado está el
Telencéfalo, que es la parte superficial, y por otro el Diencéfalo, que es la parte interna.
En cuanto al peso del cerebro, cabe decir que está estimado alrededor de los 1.380
gramos, con un margen de amplitud de 300 gramos. No está claro si existe relación
directa entre el peso del cerebro y la inteligencia, a pesar del gran número de estudios
que se desarrollan o se han desarrollado acerca del tema.
En relación con la talla, debemos decir que las personas con una gran altitud
tienen un cerebro un poco más grande que las bajas, pero con diferencias muy pequeñas
(alrededor de los veinte gramos, en proporción). En el ser humano, el cerebro ronda el
1,5 o 2% del peso corporal.
Los pliegues cerebrales son el resultado de la adaptación del cerebro a un
espacio relativamente pequeño, que es la cabeza. El perímetro craneal es algo
importante, sobre todo en recién nacidos, donde ciertas patologías cursan con aumento o
disminución de la capacidad craneal. El cerebro de un recién nacido pesa alrededor de
300 gramos, y experimentará un aumento de peso muy importante a lo largo de su vida.
Podríamos dimensionar el cerebro en unas medidas de 12x12x16, en orden de
altura, diámetro transversal, y longitud ántero-posterior.
En cuanto a su morfología, podemos señalar que superficialmente se sitúa la
corteza cerebral, que es sustancia gris. No obstante cabe señalar también que aparecen
pequeñas zonas con sustancia gris en lo que se conoce como Núcleos de la Base. Como
en todo el SNC, en el cerebro existen una serie de cavidades que reciben el nombre de
ventrículos. Dos de ellos se sitúan en el telencéfalo (ventrículos laterales) y un tercero
va a pertenecer a la parte diencefálica (es el tercer ventrículo o ventrículo medio).
ASPECTO MORFOLÓGICO
La superficie del cerebro presenta surcos y circunvoluciones. A los surcos más
llamativos les llamamos cisuras que, además de ser constantes, son más profundas y
aparecen precozmente en la escala zoológica.
Hemos de tener en cuenta que existen dos hemisferios cerebrales: uno a la
derecha y otro a la izquierda, de ahí que cuando queremos observar un hemisferio,
49
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
debamos realizar un corte sagital y medio. Existen no obstante unas estructuras que se
denominan formaciones interhemisféricas, y que ponen en contacto ambos hemisferios.
La cisura que divide los hemisferios se conoce con el nombre de Cisura
interhemisférica. Una cisura de especial interés es la Cisura de Silvio, que se bifurca en
su parte distal dando lugar a dos prolongaciones.
La tercera cisura es la de Rolando o Cisura Central, que se genera en la parte
media y se dirige de atrás hacia delante, de nuevo hacia atrás,vuelta hacia delante y
finalmente hacia atrás (cisura en zig-zag).
Una cuarta cisura sería la Perpendicular Externa, aunque también se encuentra
en la cara interna, donde se denominará Cisura Perpendicular Interna. Se encuentra en la
región posterior del cerebro.
Existen unos surcos complejos en la región cerebral posterior que se denominan cisura
Simiana (está muy desarrollada en los monos) y que delimita el lóbulo occipital (de ahí
que se denomine también cisura Preoccipital)
Podemos observar, igualmente, una cisura interna que nunca llega al borde del
hemisferio que se denomina Cisura calloso-marginal, y que delimita superiormente el
Gyrus Cinguli.
Finalmente, nos encontramos con la Cisura Calcarina, que se localiza en la parte
posterior de la cara interna del cerebro y presenta una forma característica de “acento
circunflejo”.
LÓBULOS CEREBRALES
Las cisuras que hemos observado hasta el momento, nos van a dividir el cerebro
en lóbulos. De esta manera, en la corteza cerebral vamos a poder distinguir las
siguientes regiones:
a) Lóbulo frontal: situado en la cara anterior de los hemisferios, por delante de la
cisura de Silvio y Rolando (cara externa) y rodeando la cisura calloso-marginal.
En la cara basal, se denomina lobulillo orbitario, pues se localiza por encina de
la órbita.
50
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
b) Lóbulo parietal: se observa en la cara externa e interna (no en la basal). En la
interna, se sitúa detrás de la cisura de rolando y llega hasta la cisura
perpendicular interna. En la parte inferior aparece un surco subparietal que
delimita el lóbulo. En la cara externa, lo observamos detrás de la cisura de
rolando y delante de la de Silvio.
c) Lóbulo temporal o esfeno-temporal: en la cara externa, se sitúa por detrás de la
cisura de Silvio, y llega hasta la cisura preoccipital. En la cara inferior no existe
un límite para el lóbulo, de forma que consideramos como tal el borde superior
del peñasco del temporal. En la cara interna, consideramos lóbulo temporal a la
porción de corteza que queda en la parte más baja y que llega hasta el nivel de la
cisura calcarina.
d) Lóbulo occipital: en la cara inferior, su límite es el borde superior del peñasco
del temporal, mientras que en la cara externa estará delimitado por el surco
simiano. Finalmente, se encontrará en la cara interna de los hemisferios por
debajo de la cisura perpendicular y calcarina.
e) Gyrus Cínguli o Lóbulo del cuerpo calloso: está delimitado por la cisura calloso
marginal y el surco subparietal. Rodea al cuerpo calloso.
f) Lóbulo de la Ínsula de Reil: es un lóbulo que no se ve a simple vista, pues para
su observación es necesario abrir los labios de la cisura de Silvio. Es una
estructura triangular.
FORMACIONES INTERHEMISFÉRICAS
En las zonas centrales de los hemisferios cerebrales nos vamos a encontrar una
serie de estructuras que unen los susodichos hemisferios. Son las formaciones
interhemisféricas, todas ellas compuestas por sustancia blanca (se trata de fibras que
cruzan de un hemisferio al otro).
Las formaciones interhemisféricas más importantes son las que presentamos a
continuación:
• Cuerpo Calloso: comienza en una punta y acaba en una zona redondeada.
Presenta, de esta forma, una serie de partes que reciben el nombre de pico,
rodilla, cuerpo y rodete o esplenio del cuerpo calloso. El cuerpo calloso es
cóncavo hacia arriba transversalmente, y convexa en sentido anteroposterior.
51
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Funcionalmente, el cuerpo calloso está compuesto por un conjunto de fibras
nerviosas que unen puntualmente un hemisferio con el otro. De esta manera se
pueden integrar informaciones de ambos hemisferios.
• Trígono o bóveda de los cuatro pilares: se sitúa en la parte inferior del cuerpo
calloso. Está formado por el cuerpo del trígono del que parten cuatro pilares: los
dos anteriores terminan en los tubérculos mamilares y los dos posteriores en una
región conocida con el nombre de Hipocampo.
• Septum Lucidum: es una membrana que se sitúa entre el cuerpo calloso y el
trígono.
CARA EXTERNA
CARA INTERNA
CARA INFERIOR
52
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
EL LÓBULO FRONTAL
El lóbulo frontal va a presentar una serie de surcos en su cara externa y otros en
su cara interna, que nos va a permitir dividirlo en una serie de regiones especializadas
en diversas funciones.
EL LÓBULO FRONTAL EN LA CARA EXTERNA DEL CEREBRO
En la cara externa existe un surco paralelo a la cisura de Rolando que
denominamos surco prerrolándico o precentral, así como dos surcos que parten de
forma medianamente perpendicular de dicho surco precentral y que se llaman Frontal
superior y frontal inferior. Entre el surco prerrolándico y la cisura central aparece la
circunvolución frontal ascendente (FA). Es muy importante porque en ella se sitúan
las neuronas encargadas de los movimientos voluntarios (región motora primaria). Por
delante de esta circunvolución aparecen otras tres, separadas por los surcos frontales
superior e inferior, y que se denominan frontal superior (F1) frontal media (F2) y
frontal inferior (F3). Es importante reconocer en F3 unos centros en relación con los
movimientos musculares del habla, zona descrita por primera vez por Broca (de ahí que
esta región se conozca también con el nombre de circunvolución de Broca). En la
circunvolución de Broca nos vamos a encontrar con tres partes diferentes: una de ellas
se sitúa por delante de las prolongaciones en V de la cisura de Silvio (cabeza); la
segundo es la zona del cabo, que rodea un surco medio; y la tercera parte, se denomina
pié.
EL LÓBULO FRONTAL EN LA CARA INTERNA DEL CEREBRO
En la cara interna de los hemisferios cerebrales observamos, en primer lugar, la
F1 (que se denomina frontal interna). Observamos también una parte correspondiente
con la frontal ascendente.
EL LÓBULO FRONTAL EN LA CARA INFERIOR DEL CEREBRO
En la cara inferior se observan tres surcos: un surco orbitario interno (recto) un
surco orbitario externo (curvo) y un surco en el medio que se denomina cruciforme (en
53
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
forma de H o de K). La parte más interna es la F1, y como es muy rectilínea se
denomina Gyrus Recto (o circunvolución recta). La F3 queda por fuera del surco
orbitario externo, pero se extiende por detrás del surco de Herbé (cruciforme). La F2
ocupa la superficie restante. Hay
un lugar en el que las tres
circunvoluciones coinciden, que
recibe el nombre de Cruce
Olfativo de Broca o polo frontal
funcional (en realidad la función
olfativa no es resultado del cruce
de las circunvalaciones). Este
nombre procede de la existencia
de serie de estructuras olfatorias por debajo del surco orbitario interno. Así, nos
encontramos con una dilatación, que es el bulbo olfatorio que se continúa con la cintilla
olfatoria. Ésta, en su parte terminal, se dividirá en tres raíces.
El bulbo es sustancia gris, y se continúa con la cintilla, que es sustancia blanca.
Esta cintilla se divide, muy cerca de la fisura de Silvio en una raíz interna, una externa y
otra media. Detrás de dicha división se encuentra la cisura de Silvio, y también en esta
zona encontraremos formaciones ópticas, como son la quiasma y las cintillas o
bandeletas ópticas. Queda un rombo entre cisura y cintillas que se conoce con el nombre
de Espacio Perforado Anterior, por el que pasan gran cantidad de vasos sanguíneos. A
este nivel existen también unas fibras nerviosas que lo atraviesan horizontalmente, que
reciben el nombre conjunto de Cintilla Diagonal de Foville-Broca. En la parte anterior
de la misma nos vamos a encontrar un tubérculo de sustancia gris cortical, que es el
tubérculo olfatorio. En la parte posterior de la misma nos encontramos con una zona con
muchos orificios: el verdadero espacio perforado anterior.
LÓBULO PARIETAL
En el lóbulo parietal hay un primer surco paralelo a la cisura de Rolando, que se
denomina surco poscentral o posrrolándico, y que separa una circunvolución que recibe
el nombre de Parietal Ascendente (PA). Dicha circunvolución, constituye el área
primaria de la sensibilidad. Existe un surco interparietal, que divide el lóbulo en dos
54
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
circunvoluciones: una por encima y otra por debajo, que reciben el nombre de Parietal
Superior (PS) y Parietal Inferior (PI).
La circunvolución parietal superior puede verse en la cara interna de los
hemisferios, y está limitada por un surco que recibe el nombre de surco subparietal. En
esta zona pasa a denominarse lobulillo cuadrilátero o circunvolución parietal interna.
En la circunvolución parietal inferior existen otras dos circunvoluciones
menores: una que da la vuelta a la cisura de Silvio, y otra que rodea al primer surco
temporal. Éstas se denominan circunvolución supramarginal (Silvio) y
circunvolución del Pliegue curvo, respectivamente; y están separadas por el surco
intermediario de Tensey. En esta zona se lleva a cabo la asociación significantes y
significados (conceptos e imágenes mentales), por lo que si esta área se lesiona tiene
lugar la afasia sensitiva de Wernekin (el paciente ni entiende ni habla con sentido,
articula palabras sin senso).
En el lóbulo parietal está localizada también la capacidad de conciencia del
propio cuerpo.
LÓBULO ESFENOTEMPORAL
En la cara interna, el lóbulo temporal está delimitado por una zona en la que
existe una cisura de gran importancia, que parece continuarse con la cisura calcarina
(pero nunca llegan a establecer contacto real): es la cisura del Hipocampo. Esta cisura
presenta una serie de características especiales, pues se trata de una cisura muy profunda
que se pliega sobre sí misma para no ocupar demasiado espacio.
En el lóbulo temporal podemos distinguir tres circunvoluciones, resultado de la
existencia de dos surcos temporales: T1, T2, T3. El primer surco temporal se conoce con
el nombre de surco paralelo, denominación que procede de su posición respecto a la
cisura de Silvio.
La tercera circunvolución temporal se observa también en la cara inferior, donde
estará delimitada por un surco. En la cara inferior observamos un segundo surco, que
divide el resto de la cara inferior en T4 y T5. La T5 la encontraremos también en la cara
interna.
La T4 y la T5 son circunvoluciones que se continuarán con las occipitales. Dado
que no existe una separación clara entre circunvoluciones esfenotemporales y
55
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
occipitales, los conjuntos T4/O4 y T5/O5 recibieron unas denominaciones propias
(lobulillo fusiforme y lobulillo lingual, respectivamente).
Finalmente, cabe decir que la T5 se encuentra cercana al hipocampo, de ahí que
la región que ocupa se conozca con el nombre de parahipocampo. Existe una parte del
parahipocampo que da la vuelta y forma una especia de gancho, dando lugar a una
estructura que recibe el nombre de Uncus.
La parte más alta de T5 forma parte del área de Wernike, y en él están los centros
primarios y secundarios de la audición. Finalmente, es importante porque la zona del
hipocampo tiene función directa con los procesos de memorización. También está
relacionado con el sistema instintivo primitivo y la emoción.
LÓBULO OCCIPITAL
Situándonos en la cara externa, podemos decir que el surco preoccipital de
Mayner delimita este lóbulo por su parte anterior. No obstante, existen otros dos surcos
en la cara externa del occipital, de forma que se producirán tres circunvoluciones
distintas: O1 (occipital superior), O2 (occipital media), O3 (occipital inferior).
En la cara inferior, los surcos esfenotemporales se continúan a nivel occipital, de
forma que nos encontramos con la O3, la O4 y la O5, continuando a las circunvoluciones
esfenotemporales.
La O5 “da la vuelta”, y la encontramos también en la cara interna, por debajo de
la cisura calcarina. Entre la cisura calcarina y la perpendicular interna, nos encontramos
con la última circunvolución occipital, la O6. Se trata ésta de una circunvolución que,
dado su forma triangular, se conoce como circunvolución cuneiforme o cuña.
El surco que divide la T5/O5 de T4/O4 es especialmente importante, pues marca
una huella en los ventrículos. Se le conoce con el nombre de surco colateral. Esta
característica es compartida por la cisura calcarina y la del hipocampo, que también
dejarán señal de su existencia en los ventrículos.
En el lóbulo occipital se desarrolla, como función principal, la visión. Por
consiguiente hemos de asociar los procesos y patologías visuales con localizaciones en
el lóbulo occipital.
56
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
GYRUS CINGULI
Está rodeando el cuerpo calloso, y en sus límites, se continúa con la T5, con la
que forma una cierta “unidad”. Se conocía con el nombre de limbo de los hemisferios,
de ahí la denominación del mismo como lóbulo límbico. En él residen los instintos y
las emociones y está relacionado con la función olfatoria.
Broca, describió por primera vez el lóbulo límbico, y nos dijo que podía ser
entendido como una raqueta, situada en el limbo de los hemisferios. La pala de esta
raqueta sería el parahipocampo y el gyrus cinguli, mientras que el mango sería el
sistema olfatorio. Broca no estaba en lo cierto con esta teoría, pues se demostró que por
dentro del lóbulo límbico existían unas estructuras corticales más antiguas que el propio
lóbulo, que recibieron el nombre de circunvoluciones intralímbicas. Estas
circunvoluciones intralímbicas son:
• Cuerpo abollado: situado por dentro y por encima de la cisura del hipocampo, de
sustancia gris cortical y que presenta numerosos entrantes y salidas.
• Fasciola cinérea: se trata de un tejido nervioso poco desarrollado que constituye
la continuación del cuerpo abollonado hacia el cuerpo calloso.
• Induseum Gris y Nervios de Lancisi: son unas estructuras que se localizan
pegadas a la cara convexa del cuerpo. Cuando observamos el cuerpo calloso por
encima, observamos una capa muy fina de
neuronas que se conoce con el nombre de
Induseum Gris, que estará atravesada por una
serie de fibras que van de delante atrás, que
son los nervios o tractos de Lancisi. El
induseum gris y los nervios de lancisi se
juntarán en el espacio perforado anterior,
dando lugar a la diagonal que se conoce con el
nombre de Cintilla Diagonal. Esta diagonal, mediante la cintilla del Uncus
(recorre el uncus del parahipocampo), volverá a entrar en contacto con el cuerpo
abollonado, y cerrará el sistema intralímbico.
• Núcleo amigdalino: estructura de gran relevancia situada en la profundidad de
T5.
57
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CARA EXTERNA
CARA INTERNA
CARA INFERIOR
LA ÍNSULA
Para poder observarla, es necesario abrir los bordes de la cisura de Silvio. Se
caracteriza por tener una serie de circunvoluciones muy rectas, con cuatro surcos. Uno
de ellos es más profundo que los otros y se conoce con el
nombre de surco interinsular, de forma que la ínsula
queda dividida en dos porciones: una anterior y otra
posterior. La parte anterior de la ínsula tiene tres
circunvoluciones: la I1, I2 e I3; mientras que la posterior
cuenta sólo con dos: la I4 y la I5.
No se conoce con exactitud la función de la ínsula, pero se cree que en ella se
desarrollan funciones viscerales. Lo que sí es de importancia clínica, es que la ínsula se
emplea como sistema de referencia en TAC o RMA gracias a su particular estructura
transversal.
58
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
EL CEREBRO POR DENTRO
En el cerebro, existen una serie de núcleos de materia gris que reciben el
nombre, en conjunto, de Núcleos Optoestriados. Existe un único núcleo optoestriado en
el diencéfalo de especial importancia: es el tálamo óptico. En resto de los núcleos que
estudiaremos, pertenecen a la región que conocemos con el nombre de telencéfalo, y
son:
a) Núcleo caudado
b) Núcleo lenticular.
c) Núcleo accumbens.
d) Núcleo claustrum.
e) Núcleo amigdalino.
EL TÁLAMO ÓPTICO
Es un núcleo perteneciente al diencéfalo, bastante voluminoso. Existe un tálamo
óptico en cada uno de los hemisferios cerebrales, y presenta una forma ovoidea con dos
prominencias en la parte posterior, que son los Cuerpos Geniculados (lateral y medial).
59
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Los tálamos ópticos de ambos lados se juntan por la parte de adelante y se
separan por detrás, y existe un punto en el que entran en contacto que se conoce con el
nombre de Comisura Gris o Núcleo Reuniens (aunque realmente no haya fibras que
pasen de un lado a otro). Entre los tálamos queda una cavidad que recibe el nombre de
tercer ventrículo o ventrículo medio. De esta manera, concluimos que en la pared
interna de los tálamos vamos a encontrar epitelio ependimario.
Los tálamos y el trígono entran en contacto por su parte anterior, sin embargo,
en esta localización existe una hendidura que se conoce con el nombre de agujero de
Monro, cuya función es comunicar los ventrículos laterales con el tercer ventrículo.
NÚCLEO CAUDADO
Se dice que, en conjunto, el núcleo caudado tiene la forma de un anillo
incompleto alrededor del tálamo. Se inicia en la parte anterior del mismo, siendo un
núcleo relativamente grueso (cabeza del núcleo caudado). Va disminuyendo de grosor y
se sitúa por encima, por detrás y por debajo del tálamo. La parte intermedia del núcleo
caudado se denomina cuerpo, y la región terminal del mismo, cola. Pegado a ésta, en su
zona anterior, se encuentra el núcleo amigdalino. En su inicio, las cabezas de los
núcleos caudados están muy próximas, mientras que los cuerpos se van separando, hasta
llegar a la cola, que se introduce en el lóbulo esfeno-temporal. Esta distribución del
núcleo caudado es especialmente importante, pues permiten descifrar el funcionamiento
del sistema nervioso central.
EL NÚCLEO LENTICULAR
Tiene aspecto de prisma triangular y se encuentra lateralmente al tálamo óptico.
No obstante, debemos decir que disminuye la altura a la vez que desciende, de ahí que
algunos autores lo geometricen como una pirámide triangular y no como un prisma. El
núcleo lenticular tiene dos partes muy bien diferenciadas: una externa que se conoce
con el nombre de putamen y una interna que se denomina pálidum (es más clara).
Ambas partes están separadas por una lámina medular de sustancia blanca que se
conoce con el nombre de lámina medular externa. El pálidum presenta también otra
lámina medular interna, más delgada, y que lo divide la región en pálidum medial y
lateral.
60
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El putamen y el núcleo caudado son muy parecidos histológica, anatómica y
funcionalmente, pero están separados por una serie de fibras en su mayor parte. Aún así,
juntos forman una unidad que se conoce con el nombre de Neo-estriado (o estriaculum).
El palidum, por el contrario, es una estructura totalmente diferente en su
morfología; hasta tal punto, que los anatómicos modernos hablan de un estriado dorsal,
que estaría constituido polo neoestriado y un estriado ventral, formado por el palidum y
el núcleo accumbens.
VISTA SUPERIOR
VISTA FRONTAL
61
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
FORMACIONES INTERHEMISFÉRICAS
Antes de comenzar el estudio de los ventrículos cerebrales, es de interés aclarar
ciertos aspectos sobre algunas formaciones interhemisféricas que serán de relevancia a
la hora de conocer la estructura de los distintos ventrículos.
EL CUERPO CALLOSO
Se trata de otra formación interhemisférica que permite la conexión entre las
distintas regiones de los dos hemisferios cerebrales. Como ya sabemos, el cuerpo
calloso cuenta con pico, rodilla cuerpo y
esplenio. No obstante, si observamos el
cuerpo calloso desde arriba, podemos
observar cómo las fibras transversales que
lo constituyen adquieren forma de fórceps
o tenaza en la zona anterior (lóbulos
frontales) y en la zona posterior (lóbulos occipitales), estructuras a las que se le han
dado los nombres que a continuación se exponen:
• Fórceps minor: une los polos frontales.
• Fórceps maior: une los polos occipitales. Es una estructura mayor que el
fórceps minor a causa de la mayor distancia que existe entre los polos
occipitales en relación con los frontales.
EL TRÍGONO
El trígono es una formación interhemisférica constituida por una serie de fibras
longitudinales y transversales. Las fibras longitudinales darán lugar a los cuatro
tabiques típicos del trígono, de los cuales, los dos anteriores se dirigirán a los tubérculos
mamilares y los dos posteriores a la zona de hipocampo. Los cordones de fibras que
parten del hipocampo y lo comunican con los pilares del trígono se conocen como
Fimbrias del Hipocampo (una en cada hesmisferio). Estas fimbrias suponen la
prolongación del hipocampo. De esta manera, y gracias a las fibras longitudinales, es
posible transmitir información entre el hipocampo y los tubérculos mamilares.
62
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La existencia de fibras transversales es necesaria porque es de especial
importancia que los hipocampos de ambos hemisferios estén interconectados. Gracias a
estas fibras, pues, los dos hipocampos pueden entrar en contacto.
EL SEPTUM LÚCIDUM O SEPTUM PELÚCIDUM
Se trata de una estructura que une el cuerpo calloso con el trígono, y que
presenta más altura en su parte anterior que en su parte posterior. Esta característica
permite que el Septum Lúcidum sea empleado como punto de referencia en análisis
clínicos tales como el TAC o la RMA.
VENTRÍCULOS CEREBRALES
Al igual que en cualquier parte del Sistema Nervioso Central, en el cerebro nos
vamos a encontrar con una serie de cavidades que van a recibir el nombre de
ventrículos. Específicamente existen tres ventrículos en el cerebro: ventrículo lateral
izquierdo, ventrículo lateral derecho y tercer ventrículo o ventrículo medio. A
continuación procederemos a su estudio.
63
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VENTRÍCULO MEDIO, TERCER VENTRÍCULO (DIENCEFÁLICO)
Está situado por debajo del trígono y entre los dos tálamos ópticos. Se trata de
un ventrículo aplanado en el que consideraremos dos paredes laterales, una parte
superior que se conoce con el nombre de techo, una pared anterior y otra posterior.
Como simplificación, podemos considerar al tercer ventrículo en forma de infundíbulo
(embudo).
PAREDES DEL TERCER VENTRÍCULO
Están formadas por los dos tálamos ópticos, que en su parte superior presentan el
agujero de Monro, mediante el cual se comunica el tercer ventrículo con los ventrículos
laterales. De este orificio parte un surco que llega hasta el conducto de Silvio, y que
delimita por debajo el tálamo óptico. Es el surco de Monro. La superficie que queda por
debajo del surco de Monro, pasa a denominarse Infundíbulo.
Por otra parte, están los pilares anteriores del trígono, que se dirigen a los
tubérculos mamilares pegados a la pared, pero no se observan (se incluyen dentro de la
propia pared). No obstante, estos pilares nos permiten dividir el infundíbulo en dos
partes: el subtálamo y el hipotálamo.
Con la información que poseemos, es posible ya dividir el diencéfalo en una
serie de regiones, tal y como a continuación se presenta:
a) Tálamo: constituido por los dos núcleos diencefálico.
b) Subtálamo: situado entre el surco de Monro y los pilares anteriores del trígono.
c) Hipotálamo: situado por debajo de los pilares anteriores del trígono. Situada en
esta región se localiza la glándula hipófisis.
d) Epitálamo: situado en la parte superior del tálamo. En esta zona nos
encontramos con la glándula epífisis.
Como hemos dicho ya, el tercer ventrículo se encuentra en comunicación con los
ventrículos laterales mediante los agujeros de Monro, pero también estará en contacto
con el cuarto ventrículo gracias al acueducto de Silvio.
64
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
BORDE ANTERIOR DEL TERCER VENTRÍCULO
En primera instancia, nos encontramos con el pico del cuerpo calloso, al que
llegan los pilares anteriores del trígono, que se unirán a la pared para dirigirse a los
tubérculos mamilares.
Descendiendo, observamos una estructura blanquecina que está constituida por
una serie de fibras que atraviesan el ventrículo de un lado a otro y que recibe el nombre
de Comisura Blanca Anterior. La unión de los pilares anteriores del trígono con la
comisura blanca anterior da lugar a una estructura conocida con el nombre de Bulba de
Viussens. Hacia abajo hay una lámina muy delgada, que es la lámina terminal, y que
supone la región más estrecha de las paredes del tercer ventrículo.
Un poco más caudalmente, nos encontramos con el quiasma de los nervios
ópticos y ya se observa el infundíbulo con el tallo hipofisario (el lóbulo posterior o
neurohipófisis).
BORDE POSTERIOR DEL TERCER VENTRÍCULO
En ella se distingue la parte posterior del lóbulo posterior de la hipófisis, al igual
que la región posterior del tallo hipofisario, así como una zona que protuye y que se
conoce con el nombre de Eminencia Media. Aparecen también los Tubérculos
Mamilares, y hacia la zona posterior se localiza el orificio del conducto de Silvio, que
recibe el nombre de Ano de Viussens. Existen igualmente otras fibras que pasan de un
lado al otro, formando una estructura que se denomina Comisura Blanca Posterior.
Un poco más hacia la parte superior, nos encontramos con la glándula epífisis,
que presenta forma de “piña” con una pequeña depresión anterior que se denomina hilio
de la epífisis. De ella salen unas estructuras nerviosas de forma triangular que sujetan la
epífisis y que son los Tríangulos de la Habénula. Estos triánguloS se continúan con unas
fibras blancas nerviosas que se conocen como Pedúnculos de la Habénula o Tenia
Talami.
TECHO DEL TERCER VENTRÍCULO
En el techo del tercer ventrículo nos encontramos, en un primer lugar, con una
membrana ependimaria, así como una serie de vasos que se van a introducir en esta
65
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
región. Estos vasos van acompañados de una capa de piamadre y se conocen como
plexos coroideos del tercer ventrículo. Estas estructuras que recubren el techo del tercer
ventrículo se conocen con el nombre de Membrana Tectoria. Es necesario especificar
que los vasos que recorren el techo del tercer ventrículo no llegan hasta el agujero de
Monro. Por ello, si realizamos un corte transversal por detrás de dicho agujero,
podremos observar una serie de vasos y epitelio ependimario, mientras que si el corte se
realiza a nivel del orificio de Monro, no podremos diferenciar plexos coroideos.
Es de interés señalar que el epitelio ependimario del tercer ventrículo no es igual
en todas las localizaciones, pues en ciertos emplazamientos presenta una serie de
órganos que, aunque no se sabe ciertamente su función, se sospecha que pueden estar
relacionados con la formación de LCR específicos (función endocrina). El órgano más
estudiado es el Subfornical (por debajo de los pilares anteriores del trígono). Existe una
segundo órgano que es el Paraventricular, que se encuentra en la pared del tercer
ventrículo. Finalmente observaremos un tercero bajo la comisura blanca posterior, que
se identifica como el órgano Subcomisural.
66
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VENTRÍCULOS LATERALES
Existen dos ventrículos laterales, cada uno de ellos situados en cada uno de los
hemisferios. Presentan una forma característca de “C” o herradura, con una
prolongación posterior. Esta forma es el resultado del proceso de desarrollo de los
hemisferios.
Para poder definirlo, consideraremos que el ventrículo tiene tres prolongaciones,
que parten de un punto conocido con el nombre de Cruce Ventricular. Las tres
prolongaciones se conocen con el nombre de Frontal, Occipital y Esfenotemporal. Cabe
señalar que entre las prolongaciones frontal y esfenotemporal se sitúa el tálamo óptico.
PROLONGACIÓN FRONTAL
Vamos a distinguir dos partes en dicha prolongación que van a estar delimitadas
por el agujero de Monroe. En la parte posterior aparecerá el tálamo óptico y el núcleo
caudado, mientras que en la parte anterior sólo nos encontraremos con la cabeza del
núcleo caudado.
1. Región Posterior.
Si realizamos un corte por detrás del agujero de Monro, nos vamos a encontrar
con una serie de vasos sanguíneos rodeados de piamadre que pasan por encima de los
tálamos ópticos, y que llegan, por tanto, a la base de la prolongación frontal.
Si observamos las estructuras desde un punto de vista superior, podremos
distinguir cómo la fijación de los vasos sobre los tálamos deja un surco que se conoce
67
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
con el nombre de surco coroideo. En la parte externa del tálamo aparece el núcleo
caudado. El surco que queda en la parte externa del tálamo a causa del susodicho núcleo
caudado recibe el nombre de surco opto-estriado. En él, aparecen una serie de fibras
surco
nerviosas que recorren y dan la tálamo-
estriado
vuelta al tálamo óptico,
formando una estructura que se
conoce como Taenia
Semicircular; y junto a ella,
aparece la vena optoestriada.
Ambas estructuras estarán
cubiertas por una
especialización del epitelio
ependimario que recibe el
nombre de lámina afixa.
Podemos definir, pues, la región posterior de la prolongación frontal del
ventrículo lateral (el cuerpo) como cuadrangular, con una cara superior formada por el
cuerpo calloso, con su cara interna constituida por el Septum Lucidum y su cara externa
por el núcleo caudado. La cara inferior, estaría formada por el trígono, el tálamo con sus
plexos coroideos y el surco opto-estriado (con la taenia semicircular y la vena
optoestriada). No obstante cabe decir que ciertos autores consideran el cuerpo del
ventrículo con dos caras (núcleo caudado, tálamo y trígono/cuerpo calloso) y dos
bordes.
68
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
2. Región Anterior.
A nivel anterior, desaparece el tálamo óptico y el trígono. En esta zona nos
encontraremos, pues, el cuerpo calloso y el Septum Pelucidum,
así como el núcleo caudado; de forma que en ventrículo
adquiere una forma triangular.
PROLONGACIÓN ESFENOTEMPORAL
El ventrículo lateral en esta prolongación es aplanado, un poco más pequeño que
en la prolongación frontal y presenta dos caras: una súpero-externa (cóncava hacia
adentro y convexa hacia fuera) y otra ínfero-interna (que presenta una serie de
accidentes). Por la Hendidura Cerebral de Bichat (que se establece entre el tronco
encefálico y el cerebro) llegan una serie de plexos coroideos cubiertos de piamadre
hasta el ventrículo. Adyacentes a las paredes del ventrículo existen muchas fibras
blancas que proceden fundamentalmente de la parte media del cuerpo calloso (que van
desde los lóbulos esfenotemporales de un hemisferio al otro). Estas fibras reciben el
nombre de Tapetum. Del mismo modo vamos a encontrar una parte de sustancia gris
que se va a corresponder con la cola del núcleo caudado, que como ya se ha dicho, llega
hasta el lóbulo esfenotemporal, pasando por encima del ventrículo lateral.
69
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En la cara interna, podemos observar dos huellas producidas por el surco
colateral y por la cisura del hipocampo, que van a dar como resultado la Prominencia
Colateral y el Alveus, respectivamente.
Además de lo que hemos nombrado, en esta zona vamos a encontrar una
estructura de sustancia blanca que conocemos con el nombre de Fimbria (no son más
que los pilares posteriores del trígono). Entre la propia
cisura y la fimbria, se localiza el Cuerpo Abollonado.
Finalmente, cabe decir que por delante de la
zona del núcleo caudado, se encuentra el núcleo
amigdalino. En muchas ocasiones, este último marca
también una huella en la punta del ventrículo,
característica que va a depender del paciente (esta
huella no se manifiesta en todos).
PROLONGACIÓN OCCIPITAL
Es la prolongación más pequeña de las tres. Su cara súpero-externa lisa, cóncava
hacia la parte interna y convexa hacia la externa. En ella aparecen una serie de fibras
que dan la vuelta por encima del ventrículo y llegarán hasta la cisura calcarina. Son las
radiaciones ópticas.
70
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En la cara ínfero-interna nos vamos a encontrar una marca producida por la
cisura calcarina que se conoce con el nombre de Espolón de Morano (Ergot), así como
una segunda huella producida por el surco colateral que se denomina Eminencia
Colateral. Finalmente, encontramos una última impresión en el techo de la prolongación
occipital que es el resultado del paso de fibras del fórceps maior del cuerpo calloso.
La prolongación occipital carece de plexos coroideos, pues éstos se sitúan
únicamente en las prolongaciones frontal y esfenotemporal, así como en el cruce
ventricular.
ZONA DEL CRUCE
Se encuentra en la parte posterior del tálamo y en ella se juntan las tres
prolongaciones ventriculares. Tiene plexos coroideos que llegan por su cara interna.
Una vez conocidas las tres prolongaciones y la zona del cruce, podemos
entender cómo se distribuyen los plexos coroideos en el conjunto de los ventrículos
laterales. De esta manera, observamos que siempre proceden de la parte interna y que
nunca llegan a la prolongación occipital ni más allá del agujero de
Monro. Resultaría, pues, una estructura semejante a la que podemos
ver en el esquema de la izquierda.
GENERALIDADES SOBRE ESTRUCTURAS Y COMPONENTES NERVIOSOS
A lo largo de nuestro estudio, hemos hecho continua referencia a una serie de
elementos y estructuras directamente relacionados con el Sistema Nervioso. A
continuación, se pretende aclarar estos conceptos.
EL LÍQUIDO CEFALORRAQUÍDEO O LIQUOR CEREBROESPINALIS
El LCR es un fluido totalmente incoloro, por lo que se dice que tiene el color de
agua o cristal de roca. Es absolutamente transparente. Si esta característica cambia, nos
encontramos ante un LCR patológico.
En cuanto a su localización, cabe decir que El LCR se sitúa dentro y fuera (en
los espacios meníngeos y subaracnoideos) del sistema nervioso central. Se calcula que,
71
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
en un adulto normal, existe un promedio de 150 cm3 de líquido cefalorraquídeo, de los
cuales, alrededor 100 cm3 se localizan en los espacios leptomeníngeos y 50 cm3 en las
cavidades del SNC.
El líquido cefalorraquídeo se caracteriza porque, en el organismo, se rige por un
proceso que se inicia con su formación, se continúa con su circulación y finaliza con su
reabsorción; tal y como a continuación se explica:
a) Formación: el LCR se forma diariamente en cantidades que rondan los 500 cm3
(se fabrica constantemente, como todos los fluidos orgánicos). Su formación se
lleva a cabo dentro de las cavidades del SNC (el 90 %), la mayor parte en los
plexos coroideos de los ventrículos, principalmente en los pertenecientes a los
ventrículos laterales. Existe una pequeña parte, no obstante, que se puede formar
en emplazamientos donde existan capilares, pero es una cantidad poco
significativa. Su constitución se realiza en parte por secreción y en parte por
paso directo de sustancias a través de la membrana (depende del grosor de las
moléculas).
b) Circulación: el LCR está circulando de
forma continua por el interior de los
ventrículos. Posteriormente, sale al
exterior por los orificios de Luschka y
Magendie, y circula por los espacios
leptomeníngeos hasta que se reabsorbe.
c) Reabsorción: tiene lugar en las vellosidades de Pachioni, estructuras situadas en
las leptomeninges (principalmente en las blandas). De esta manera, se puede
comprobar cómo la mayor parte del LCR se reabsorbe fuera del SNC (alrededor
del 90 %).
A causa de este proceso al que está sometido el LCR, existe una patología propia
del mismo en la que se forma pero no se reabsorbe,
dando lugar a un aumento de la cantidad de LCR en el
sistema nervioso, una enfermedad conocida con el
nombre de Hidrocefalia. Cabe decir, no obstante, que
cualquier obstrucción en la circulación del LCR, o
cualquier tipo de fallo en la formación y reabsorción
puede acabar produciendo una hidrocefalia. En estos
72
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
casos, se produce un aumento de líquido en la cavidad craneal. Debido a ello, pueden
ocurrir dos cosas:
• En un recién nacido, la cavidad craneal aumenta considerablemente (de ahí
que sea especialmente importante medir el perímetro craneal a los recién
nacidos).
• En un adulto, el aumento de líquido provoca una presionando en el SNC que
va en aumento y que provoca que el cerebro se vaya atrofiando
paulatinamente (los pacientes pierden el equilibrio, movilidad, pueden
padecer demencia,….)
Otro aspecto de importancia es la presión del LCR, que varía según el punto del
SNC donde se mida y según la posición del paciente. Así, por ejemplo, una persona en
sedestación presenta una presión media del LCR que ronda los 25 cm de H2O. Por el
contrario, en una persona en decúbito, esta presión desciende aproximadamente hasta
los 10 cm de H2O. En bipedestación, los valores de presión pueden llegar a cero e
incluso a valores negativos.
LAS MENINGES
Las meninges que nos podemos encontrar en el cerebro son las tres clásicas que
ya había enumerado Bichat en sus estudios: la duramadre, la piamadre y la aracnoides.
• Duramadre: tapiza todos los agujeros del interior del cráneo, rodea todo el
SNC y va a mezclar fibras con todos los elementos que entran en el cráneo.
Como la mayor parte de los agujeros del cráneo se sitúan a nivel de la base,
podemos decir que la duramadre se pega más a la base que a la bóveda (algo
que tendrá especial relevancia clínica). La duramadre también envía tabiques
divisorios.
73
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Piamadre o lámina portadora de vasos: todos los vasos que se introduzcan en
el sistema nervioso irán acompañados de ella. Va a tapizar también todas las
superficies del sistema nervioso, de manera que cualquier zona superficial
del SNC va a ir acompañada de una capa de piamadre.
• Aracnoides: presenta dos partes. Por un lado, encontramos una lámina que se
pega a la duramadre y por otra una serie de trabéculas fibrosas que se pegan
a la piamadre y que parten de la primera lámina. Por medio de estas
trabéculas quedan los espacios subaracnoideos (lugar donde se encuentra el
LCR).
Una particularidad a comentar acerca de las meninges es que existe un par
craneal, más concretamente el nervio óptico, que va también rodeado por meninges y
LCR, pues, aunque es considerado como un nervio, en realidad constituye una parte del
SNC.
LA DURAMADRE
Como ya hemos dicho, la duramadre es una de las tres meninges que recubren el
cerebro (así como todo el SNC). También hemos comentado que se encuentra más
fijada a la base del cráneo, pero hemos de especificar que, en la zona sagital y media de
la calota craneal, la duramadre se adhiere firmemente a la misma. Además, es de interés
señalar que la duramadre craneal tiene capacidad osteogénica en su parte más externa,
pues presenta osteoblastos a su estudio con microscopía.
74
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Del mismo modo, cabe señalar que en la bóveda del cráneo podemos delimitar
una zona que se conoce con el nombre de cuadrilátero de Gerard Marchand. Se trata de
una región donde se producen la gran mayoria de los hematomas y hemorragias
epidurales. Sus límites están definidos por una línea vertical que viene por el borde de
las alas menores del esfenoides y llega hasta la rama ascendente del malar en la parte
anterior, por línea horizontal que viene dada por el arco cigomático a nivel inferior y,
finalmente, por dos bordes curvos que se identifican con la zona parietal.
Las hemorragias epidurales se producen entre el hueso y la duramadre (espacio
epidural).
Es de especial interés señalar que el interior del cráneo esta compartimentalizado
parcialmente por una serie de tabiques de la duramadre.
Estos tabiques reciben el nombre de Hoz del Cerebro,
Hoz del Cerebelo y Tienda del cerebelo. La hoz del
cerebro y la del cerebelo forman un tabique vertical,
mientras que la tienda constituye un tabique horizontal.
Se trata de tabiques incompletos que dejan un orificio con
forma oval y de gran magnitud entre ellos para permitir que el tronco encefálico se
conecte con el cerebro. Es el Agujero Oval de Pachioni (por el agujero de Pachioni sólo
entra la parte del tronco encefálico perteneciente al mesencéfalo).
75
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La tienda separa el cerebro del cerebelo, de forma que existe una separación de
material fibroso procedente de duramadre entre ellos.
Los tabiques que hemos estudiado tienen relevancia en patologías, como es el
caso de los traumatismos
craneoencefálicos o aumento de
alguno de los compartimentos que
presiona las estructuras hacia el lado
contrario. Por ejemplo, cuando se
produce un hematoma epidural por
encima de la tienda, pueden
provocarse hernias del lóbulo
temporal, o un enclavamiento del
mismo a causa de la presión ejercida
por el susodicho hematoma.
LA HOZ DEL CEREBRO
Como ya sabemos, la hoz del cerebro es una estructura derivada de la duramadre
que compartimentaliza la cavidad craneal y que se sitúa entre los dos hemisferios. Para
estudiarla, hemos de decir que presenta dos caras laterales planas, tres bordes (convexo
o adherente, cóncavo y base) y un vértice que se va a fijar en la apófisis Crista Galli del
etmoides.
SENOS VENOSOS DE LA DURAMADRE
A nivel del interior del cráneo, el sistema venoso debe asegurarse la salida de la
sangre independientemente de la posición en la que el individuo se encuentre. Por ello,
existen una serie de espacios y conductos que se mantienen abiertos siempre a causa de
su rigidez, y que permiten la salida de sangre. Son espacios que quedan entre los
tabiques de duramadre que se van a llenar de sangre venosa y que van a recibir el
nombre de senos venosos. Los más importantes son:
76
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
a) Seno longitudinal superior: discurre por el borde superior o convexo de la
hoz del cerebro y puede presentar una prolongación (que no es constante)
que se introduce por el agujero etmoido-frontal y establece una
comunicación entre el drenaje venoso de las fosas nasales y el seno
longitudinal superior. Esta conexión, en caso de existir, se realiza mediante
la vena de Sabatier y Blandin. El seno longitudinal cobra especial relevancia
en los recién nacidos, pues en el inicio del desarrollo cuando las fontaneros
todavía no se han osificado, es posible acceder a él mediante la fontanela
bregmática bregmática (la que se establece entre el hueso frontal y los
parietales) para realizar un drenaje en caso de aumento de la presión venosa.
No obstante, esta técnica sólo debe ser usada en casos de extrema urgencia,
pues se corre el riesgo de producir una tromboflebitis en el seno longitudinal
del paciente.
a) Seno petroso superior: seno que discurre por la tienda del cerebelo, en
sentido horizontal. Va a desembocar en otro seno mucho más grueso que él,
que se conocerá con el nombre de seno lateral.
b) Seno lateral o sigmoideo: recorre también el borde adherente de la tienda y
se va a continuar con la yugular interna a través de una dilatación que deja
esta última a su entrada en el cráneo y que se conoce con el nombre de golfo
de la yugular.
c) Seno recto: se sitúa en la base de la hoz del cerebro y es muy rectilíneo. En
él va a desembocar el seno longitudinal inferior, además de una vena corta y
muy ancha, que se conoce con el nombre de Ampolla o Gran vena de
Galeno, a la que llegan numerosas venas cerebrales internas.
d) Seno longitudinal inferior: recorre el borde cóncavo de la hoz del cerebro. Se
trata de un seno de pequeñas dimensiones que termina en el seno recto.
e) Seno occipital posterior: discurre por el borde adherente de la hoz del
cerebelo.
Los senos venosos que hemos visto hasta el momento, se unen en una
anastomosis que se sitúa a nivel de la protuberancia occipital y que se conoce con el
nombre de Prensa de Herófilo. Habitualmente, en posición anatómica, la sangre llega
desde el seno longitudinal superior hasta los senos laterales, que la drenan a la yugular.
77
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Además de estos senos que hemos nombrado, existen otros muchos que se sitúan
en emplazamientos independientes de los tabiques de duramadre, como es el caso de los
siguientes: seno cavernoso, el seno de Berschef o esfenoidal, otro alrededor de la
hipófisis, que es el seno coronario de Ridley.
1) Seno cavernoso: se sitúa a ambos lados de la silla turca. La duramadre llega
hasta la parte superior de la hipófisis, donde hace una forma de techo que se
denomina tienda y que tiene un agujero central que deja pasar el tallo. A ambos
lados de la hipófisis, la duramadre se desdobla dando lugar a unas cavidades que
se van a llenar de sangre y que constituirán senos cavernosos. Tienen gran
importancia desde el punto de vista médico por el gran número de nervios y
vasos que hay en este nivel, así como por el hecho de que dichos senos van a
estar comunicados con las venas de la cara y con las venas del interior del
cráneo, por lo que una bacteria podría llegar al endocráneo a través de estas
venas. Esta posibilidad, hizo que los antiguos anatómicos denominaran la red
venosa que va desde la nariz a los laterales de la boca como El triángulo de la
Muerte. Los elementos vásculo-nerviosos que se sitúan en el seno cavernoso
78
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
son, por un lado, la carótida interna, que se sitúa dentro del propio seno y va a
ir rodeada de trabéculas fibrosas dependientes de la duramadre que la van a
sujetar y que se conocen como Cuerdas de Willis. Existe también un par craneal
que se introduce en el interior del seno cavernoso: es el VI par o Motor Ocular
Externo, que va siempre por fuera de la carótida interna, acompañado de una
cubierta meníngea y sujeto al seno por una serie de cordones fibrosos al seno.
De igual modo, por la pared del seno cavernoso van a situarse tres nuevos pares
craneales, que son la rama oftálmica del trigémino (V par), el IV par y el III.
2) Seno coronario de Ridley: se sitúa alrededor de la tienda de la hipófisis, dando
lugar a una especie de corona que rodea su tallo.
LAS GRANULACIONES DE PACHIONI
Son estructuras meníngeas, más concretamente de aracnoides, que se introducen
como espinas a nivel de la duramadre y del hueso. En la superficie de estas
granulaciones queda una fina capa de duramadre que no impide el intercambio de
sustancias. Cuando se
introduce en el hueso, suele
dar lugar a lagunas venosas.
Gracias a las granulaciones
de Pachioni, el Líquido
Cefalorraquídeo puede ser
reabsorbido, bien en las
lagunas del hueso o en los
senos venosos.
79
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El sistema nervioso va a desarrollarse a partir de la capa más externa del
embrión, que es el Ectodermo, a partir de una serie de procesos que implican la
formación, en primer lugar, de la Placa Neural, que crece y se pliega dando lugar al
Canal Neural. Finalmente, este canal se cerrará generando el Tubo Neural.
Todo este proceso sólo es
posible gracias a la existencia de una
estructura que se encuentra en el
embrión trilaminar y que es la
notocorda. Se trata de la estructura
promotora del crecimiento de la placa
neural y por consiguiente, de los
siguientes pasos del desarrollo del
sistema nervioso.
Previa formación del tubo neural, los bordes que forman el canal neural
adquieren especial importancia y dan lugar a la cresta neural. La importancia de esta
estructura reside en que las células que la forman tienen gran capacidad de migracón y
se van a extender por muchas zonas del organismo.
Para la constitución del tubo neural, los bordes del canal empiezan a soldarse en
la zona media (lo que será la zona dorsal de la nuca), y se van fusionando hasta dejar
únicamente dos orificios en los extremos: el Neuroporo Anterior y el Neuroporo
Posterior. Estos orificios se cerrarán en momentos distintos del desarrollo: el neuroporo
anterior se va a cerrar el día 25 después de fecundación, mientras que el posterior
tardará dos días más. Finalmente, cuando el tubo neural está ya formado, va quedar por
debajo de las células de la cresta y el ectodermo (que formará la futura piel).
80
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
ESTRUCTURA DEL TUBO NEURAL
Si observamos el tubo neural en un corte transversal, podemos distinguir un
surco que se conoce con el nombre de Surco Limitante de Hiss, que se origina como
resultado de un crecimiento exponencial de neuronas por delante y por detrás de la zona
que va a constituir el surco. Las neuronas que
crecen por delante del mismo van a ser
principalmente neuronas motoras y las que
crecen por detrás van a estar relacionadas con
funciones sensitivas. Dadas las distintas
funciones que desempeñan, las neuronas que
quedan por delante del surco forman lo que se
conoce como Placa Basal, mientras que las que
quedan por detrás constituyen la Placa Alar. Las
regiones celulares más delgadas que quedan en los bordes ventral y dorsal se conocen
con el nombre de Placa Tectriz (o del techo) y Placa Solar (o del suelo),
respectivamente.
Por otra, podemos observar que las paredes del tubo se estructuran formando tres
capas, que son las que a continuación presentamos:
1. Capa neuroepitelial: es la más cercana a la luz del tubo.
2. Manto: es la capa celular intermedia.
3. Capa marginal: es la más externas de las capas que constituyen la pared del tubo
neural, y por fuera de ella se encuentra una cubierta de piamadre primitiva.
En un primer momento, existe una única capa, la neuroepitelial. Pero las células
de esta capa envían una serie prolongaciones hasta la perifería, donde se encuentra la
piamadre primitiva y, aprovechando esta prolongación, las células empiezan a subir, y
llegan a la parte más periférica, donde se dividen en dos. Cuando esto sucede, se
originan dos células, de las cuales una vuelve a descender empleando la prolongación
como guía y la otra se desprende. De esta manera, las tres capas van a estar formadas
por una única, por lo que se dice que es una capa seudoestratificada. El resultado de
todo el proceso que hemos explicado es que, mientras que unas células quedan
atrapadas, otras se sueltan. Las células liberadas van a dar lugar a dos estirpes celulares,
que son:
81
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Neuroblastos: precursores de neuronas.
• Espongioblastos: precursores de células de la gliales.
Esta repartición en capas de las paredes del tubo van a permanecer sin variación
en la médula espinal (donde se encuentra la sustancia blanca en la perifería y la gris en
el centro), mientras que en el encéfalo, por el contrario, las células del manto van a
emigrar utilizando las prolongaciones hacia la capa marginal, por ello la sustancia gris
queda en la perifería y la blanca en el centro (como ocurre en cerebro y cerebelo). Estos
aspectos son especialmente importantes para el estudio de la etiología de ciertas
enfermedades del sistema nervioso que se cree son resultado de errores en este proceso
de emigración.
HISTOGENIA DE LAS CÉLULAS DEL SISTEMA NERVIOSO CENTRAL
A partir de las células que se liberan en el proceso de formación de las capas de las
paredes del tubo neural se van a originar dos grandes líneas celulares: el neuroblasto y
el espongioblasto (o glioblasto). Los neuroblastos terminarán su proceso de evolución
en neuronas, mientras que los glioblastos darán lugar a astocitos u oligodendrocitos.
Hoy día es de relevancia el estudio de la génesis de estas células, pues se considera que
82
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
sería posible la formación de nuevas neuronas a partir de sus precursoras. Para ello se
lleva a cabo un profundo análisis de las cascadas de señalización que activan el proceso
de diferenciación y se desarrollan métodos de localización de las células precursoras.
Hasta el momento conocemos más de cincuenta factores que dan lugar a la formación
de neuronas.
Los distintos pasos por los que debe pasar un neuroblasto apolar (sin
prolongaciones) para llegar a convertirse en una neurona se denominan neuroblasto
bipolar y neuroblasto unipolar.
83
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Los glioblastos van a dar, en primera instancia, astroblastos u oligodendroblastos
y, a partir de ellos, las células constitutivas de la glía (sólo dentro del sistema nervioso
central, pues las células gliales del sistema nervioso periférico las forman las células
procedentes de la cresta neural).
Otra línea celular la constituyen aquellas células que quedan formando parte de
los Epitelios Ependimarios, y que son las células primitivas que quedan en la capa
neuroepitelial). Se piensa que entre ellas pueden quedar células que, siendo estimuladas,
pueden dar lugar a neuronas.
LA CRESTA NEURAL
Las células de la cresta neural son las que van a dar lugar a las células que
estarán situadas fuera del sistema nervioso, aunque estén en relación con él. Por
ejemplo, a partir de ella se generarán las neuronas sensitivas de los ganglios, todas las
células vegetativas del sistema nervioso periférico (incluyendo la médula de la glándula
suprarrenal), el sistema cromafín del aparato digestivo (incluso los ganglios vegetativos
que están en relación con él),… Las leptomeninges (piamadre y aracnoides) también
provienen de la cresta neural, a diferencia de la duramadre, que es mesodérmica.
Hemos de señalar también que existe algún mesodermo que se origina a partir de la
cresta neural, y que se conoce con el nombre de Ectomesénquima. Los arcos
branquiales son un ejemplo de estructuras derivadas del ectomesénquima. Otro ejemplo
que podemos nombrar son son los odontoblastos, que dan lugar a la dentina y también
proceden del ectomesénquima.
FORMACIÓN DEL ENCÉFALO
El tubo neural empieza a crecer exponencialmente
en la región donde se situará el futuro encéfalo. De esta
forma se forman tres vesículas, que se conocen como
Prosencéfalo (la superior), el Mesencéfalo (la media) y el
Romboencéfalo (la inferior). Además, el tubo neural
empieza a plegarse, por una lado en la región del
mesencéfalo por otro en la zona final del romboencéfalo.
84
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Del prosencéfalo parte una placoda óptica y después una vesícula óptica que
darán lugar al globo ocular. Con esto, podemos entender porqué el nervio óptico no es
un nervio periférico “normal”, sino que forma parte del SNC y va a estar envuelto por
cubiertas meníngeas. Lo mismo va a suceder con el oído y el nervio estatoacústico.
Las tres vesículas de las que antes hemos hablado van a seguir evolucionando,
de forma que algunas de ellas se dividirán, dando lugar a un embrión que va a presentar
cinco vesículas distintas, expuestas a continuación:
• Del prosencéfalo van a surgir dos nuevas vesículas, que se conoceran como
Vesícula Telencefálica (que va a dar lugar al telencéfalo) y Vesícula
Diencefálica (que dará lugar al
diencéfalo).
• El mesencéfalo no se va a dividir,
y permanecerá como Vesícula
Mesencefálica (que originará el
mesencéfalo).
• El romboencéfalo, al igual que el
prosencéfalo, va a dividirse en dos
vesículas que recibirán el nombre
de Metencéfalo (precursor del
bulbo y la protuberancia) y
Mielencéfalo (a partir del cual se
generará el bulbo raquídeo).
85
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Durante las siguientes semanas del desarrollo, el telencéfalo empieza a crecer
mucho lateralmente, dando lugar a dos vesículas que se proyectarán y crecerán hacia
atrás, llegando a cubrir el diencéfalo y constituyendo, de esta manera, el cerebro. En un
primer momento, el cerebro es liso, pero a medida que el telencéfalo va creciendo, se va
plegando, y en los últimos estadíos de vida intrauterina ya se identifican casi todos los
surcos y circunvoluciones de un cerebro desarrollado.
CRONOLOGÍA DEL DESARROLLO DEL SISTEMA NERVIOSO
El sistema nervioso comienza a desarrollarse a las tres semanas después de la
fecundación. Comienza a formarse el canal y el tubo neural, dejando los neuroporos que
acabarán por cerrarse el día 25 y el 27. Posteriormente, se va a desarrollar el encéfalo,
con una cronología que se establece entre los seis y los nueve meses.
PATOLOGÍAS CAUSADAS POR FALLOS EN EL DESARROLLO DEL SNC
Los errores en el desarrollo del sistema nervioso central suelen darse en los
primeros estadíos del mismo y, cuanto más precozmente ocurran estos fallos, más
problemas causarán. Algunas de las patologías más comunes son:
• Espina bífida.
• Hidrocefalia.
• Malformación del Arnold-Chiari.
86
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
ESPINA BÍFIDA
Existen diversos tipos de espina bífida, que implican distintos grados de
gravedad. Algunas de ellas son las que presentamos a continuación:
• Espina bífida oculta: alrededor de la médula se formarán las vértebras. Uno de
los fallos más comunes (1:100) es que no se forme el arco vertebral. Su
diagnóstico suele ser casual, ya que suele ser asintomática. Puede localizarse en
cualquier región de la columna pero es frecuente en la zona lumbar. En
ocasiones, en el lugar donde se sitúa la espina bífida, aparecen unos pelillos que
pueden ser indicio de su existencia.
• Meningocele: existe un tipo de espina bífida en la que las meninges protuyen al
exterior, dando lugar a un bulto formado por duramadre y LCR. Tampoco se
forma el arco vertebral. El bulto que se forma puede ser un quiste (que no
comunica la médula con el exterior) o a una fístula (que sí comunica con la
médula espinal). Al igual que las meningoceles, pueden producirse
encefaloceles.
• Mielomeningocele: en este caso, además de las meninges y el LCR, va a protuír
también un poco la médula espinal.
• Espina bífida grave: existen casos en los que la médula no llega a desarrollarse,
quedando en forma de placa neural.
• Anencefalia: en los pacientes anencefálicos no se forma el encéfalo, o está muy
poco desarrollado. No son individuos viables.
87
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
HIDROCEFALIA
La hidocefalia consiste en un aumento del volumen de los ventrículos a causa de
una acumulación de LCR. Este aumento de líquido cefalorraquídeo provoca un aumento
de presión en el encéfalo.
Algunas hidrocefalias que son congénitas,
y la obstructiva es la más frecuente. En ella, la
acumulación del LCR es el resultado de una
obstrucción en su proceso de reabsorción.
La hidrocefalia es muy común en recién nacidos, de ahí la importancia de medir
el perímetro craneal en los reconocimientos pediátricos.
MALFORMACIÓN DE ARNOLD-CHIARI
Se trata malformación del sistema nervioso en la que parte del bulbo y el
cerebelo se introduce en la columna vertebral. Es frecuente, y debe ser identificada
rápidamente, porque puede traer consecuencias neurológicas a largo plazo.
88
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Para poder comprender la complejidad de las estructuras encargadas de la
vascularización del encéfalo, es preciso que antes se clarifiquen algunas de las
cuestiones que la caracterizan. De esta manera, podemos enumerar las siguientes
características acerca de la vascularización y circulación venosa en el encéfalo:
• La irrigación encefálica carece de vasos linfáticos, pero si posee arterias
y venas propias.
• El flujo sanguíneo total es de 800 ml por minuto.
• El flujo por cada 100 gramos de tejido cerebral está cuantificado,
aproximadamente, en 50 ml/min.
• El tiempo circulatorio ronda los siete segundos.
• El encéfalo humano emplea el 20 % de la sangre bombeada por el
corazón.
• El encéfalo va a consumir el 20 % de la glucosa y el 20 % del oxígeno
del total del organismo
La dependencia del encéfalo de la circulación sanguínea para la obtención de
glucosa y oxígeno es enorme. De hecho, si se priva al encéfalo del aporte sanguíneo
durante determinado tiempo, se producen una serie de procesos y secuelas que
resumimos a continuación:
• 10 segundos de isquemia llevan a una pérdida de conciencia.
• 20 segundos de isquemia provocan el cese de la actividad bioeléctrica
cerebral.
• 5 minutos de isquemia van a provocar un daño irreversible en el encéfalo.
• 10 minutos de isquemia producirá la muerte cerebral.
PARTICULARIDADES DE LA CIRCULACIÓN ENCEFÁLICA
Dadas las características y localización del encéfalo, la circulación que permite
su irrigación va a presentar una serie de particularidades específicas, tal y como a
continuación presentando:
• El encéfalo está situado en un compartimento cerrado e inextensible, por lo que
la toda la sangre que penetra en el endocráneo debe salir de él. Cualquier
89
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
obstrucción en la entrada o la salida, va a provocar problemas graves, como
pueden ser, por ejemplo, hematomas.
• Los vasos se hallan sometidos mecánicamente a presiones y contrapresiones
(presión intramural).
• El ser humano es bípedo, por lo que la morfología, la hemodinámica y la
regulación es específica.
• Las neuronas son irremplazables, así como tremendamente dependientes del
oxígeno y la glucosa.
• Existen mecanismos compensatorios que impiden el déficit circulatorio.
• La circulación intracerebral es terminal (en exterior todo se anastomosa, en el
interior, nada se anastomosa).
ARTERIAS QUE IRRIGAN EL CEREBRO
Al igual que todos los aspectos de la circulación encefálica, las arterias
cerebrales van a tener una serie de características definitorias. Las más importantes son
las que ahora enumeramos:
• Poseen un trayecto frecuentemente variable e incluso asimétrico.
• Presentan un territorio de distribución fijo.
• Se distribuyen por cisuras o surcos.
• Su segmento inicial es curvilíneo o rectilíneo. Las arterias cerebrales son, en su
conjunto, curvas. Esta curvatura será variable a lo largo de la vida del individuo,
y será directamente proporcional a la edad.
• Sus ramas de distribución son frecuentemente sinuosas, hasta el punto de que su
longitud es el triple de lo que correspondería en relación con su trayecto (son
muy curvas).
• Suelen encontrarse por debajo de la aracnoides, recubiertas de piamadre.
VASCULARIZACIÓN ARTERIAL DEL ENCÉFALO
El primero de los aspectos a tratar en lo que se refiere a la vascularización
arterial del encéfalo, es que la sangre llega al mismo a partir de cuatro troncos arteriales.
90
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Estas arterias son dos carótidas internas (la derecha y la izquierda) y dos arterias
vertebrales (derecha e izquierda).
Como norma general, hemos dicho que las anastomosis que se establecían en el
Sistema Nervioso se localizaban en la perifería y no en el interior de los tejidos,
formando así las anastomosis corticales. Lo que no se ha dicho hasta el momento es
que, a nivel encefálico, podemos encontrarnos con tres tipos de anastomosis:
a) Extracraneales: son muy poco importantes y carecen de valor funcional
relevante.
b) Polígono arterial de Willis: es tremendamente importante y permite que las
cuatro arterias se anastomosen en la base del cráneo, de modo que algún
fallo en una de ellas puede ser compensada por las otras.
En cuanto a la irrigación del encéfalo en su conjunto, hemos de decir que se
distinguen cuatro sistemas arteriales que llevan a cabo esta función. Son los siguientes:
a) Cortical o periférico: correspondiente a la corteza.
b) Central para núcleos grises: vascularizan núcleos de sustancia gris
intracerebrales.
c) Ventricular o coroideo: a partir de una serie de arterias que luego
trataremos se forman los plexos coroideos para los tres primeros
ventrículos.
d) Basal: se encarga de llevar sangre a la base del encéfalo).
VASCULARIZACIÓN ARTERIAL DEL ENCÉFALO
Una vez comprendida en su conjunto la circulación encefálica vamos a
comenzar con el estudio pormenorizado de las arterias más importantes que llevan a
cabo esta función.
LA CARÓTIDA INTERNA
Como primer aspecto destacable, cabe decir que las carótidas derecha e
izquierda nacen de forma diferente. La carótida primitiva derecha parte del tronco
braquiocefálico, mientras que la carótida primitiva izquierda lo hace directamente del
cayado de la aorta.
91
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La carótida primitiva se divide en dos ramas: una que sigue hacia el encéfalo
(carótida interna) y otra que se dirige hacia las envueltas del cráneo (carótida externa),
como es la cara o el cuello.
La carótida interna va a penetra en el endocráneo por el agujero carotídeo, que
se encuentra en el peñasco del temporal. Se introduce dentro de él desde la base y hace
un codo. El agujero carotídeo se encuentra en el borde del peñasco, de forma que
cuando la arteria sale por
dicho orificio, se encuentra
con el seno cavernoso, en el
que se introduce y por el que
discurre hasta su sección
terminal. Dentro del seno
cavernoso, la carótida va a dar
lugar a una forma de “S” que
se conoce con el nombre de
Sifón Carotídeo. Se trata de
una región de especial
relevancia clínica, pues en
ella son frecuentes las
dilataciones aneurismáticas.
Como toda arteria, la carótida va a dar una serie de ramas terminales que son las
que a continuación se definen:
1. Arteria oftálmica: se va a dirigir hacia la zona del ojo. Algunos autores
consideran esta arteria como una rama colateral y no terminal de carótida, pues
parte un poco antes de la división de la misma en las otras cuatro ramas.
2. Arteria Cerebral Anterior: se dirige a la cara interna de los hemisferios.
3. Arteria Silviana o Cerebral Media: va a introducirse por la cisura de Silvio para
llegar hasta la cara externa de los hemisferios.
4. Arteria Coroidea Anterior: se dirige a los plexos coroideos y va a formar parte
de ellos. Es la arteria más pequeña.
5. Arteria Comunicante Posterior: se trata de una arteria que forma parte del
polígono de Willis.
92
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
LA ARTERIA SUBCLAVIA
La arteria subclavia derecha parte también del tronco braquiocefálico y va a dar
rápidamente una rama que constituye la arteria vertebral. La izquierda partirá
directamente del cayado de la aorta, pero también dará una arteria vertebral como rama
colateral. Estas arterias vertebrales van a dirigirse hacia la apófisis transversa de las
vértebras y pasarán por los agujeros transversos de las seis primeras cervicales,
acompañadas de un plexo venoso y el nervio de François-Frank. En el atlas, las arterias
vertebrales pasan por detrás de las masas laterales y se introducen en el conducto
raquídeo, de forma que penetran en el endocráneo.
Un hecho de importancia clínica es que la arteria vertebral se encuentra cerca del
cuerpo de las vértebras. Por ello cuando se produce artrosis en los cuerpos, pueden
producirse problemas más o menos graves en las paredes de las susodichas arterias
vertebrales, como es el caso de vasoconstricción de las misas.
93
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Una vez que las arterias vertebrales se introducen en el cráneo, se unen dando
lugar al Tronco Basilar, por ello al conjunto se le conoce con el nombre de Sistema
Vértebro-basilar. El tronco basilar va a dividirse, a su vez, en dos arterias terminales
que recibirán el nombre de Arterias Cerebrales Posteriores.
Las arterias vértebro-basilares y las arterias carótidas internas no presentan
anastomosis extracraneales que presenten una relevancia particular.
ARTERIAS QUE VASCULARIZAN EL TRONCO DEL ENCÉFALO
En el sistema nervioso, a nivel del bulbo y el puente, las arterias se sitúan en la
perifería y posteriormente dirigen sus ramas hacia el interior; tal y como ocurría en la
médula espinal. Las arterias que vascularizan el tronco del encéfalo provienen, en su
mayoría, de la parte anterior del mismo; y las clasificamos en tres grupos:
a) Arterias Paramedianas: proceden de la zona anterior y vascularizan la zona
sagital y media, por lo que su longitud es pequeña. Parten del tronco basilar y se
dirigen directamente hacia bulbo, puente y mesencéfalo. Van a denominarse en
función de la región a la que irrigen (por ejemplo, diremos que el bulbo raquídeo
está inervado por las arterias paramedianas del bulbo).
b) Arterias Circunferenciales Cortas: dan la vuelta al tronco encéfalo y vascularizan
la cara lateral del encéfalo. Las arterias circunferenciales parten de las arterias
vertebrales o del tronco basilar. Existen también una serie de arterias radiculares
que van a vascularizar los pares craneales. Algunas de estas arterias adquieren
relevancia porque van a dar lugar a ciertas patologías en los humanos. Es el
caso, por ejemplo, de la arteria de la fosilla lateral del bulbo, que se lesiona en
ocasiones, dando un síndrome clínico característico. La arteria de la fosilla
lateral del bulbo suele dar varias ramas para los nervios mixtos (IX y X par), así
como una rama que se dirige a la fosilla supraolivar. Otra ejemplo de arteria
circunferencial corta es la que se dirige e irriga al trigémino.
c) Arterias circunferenciales largas: dan la vuelta completa al tronco encefálico y
van a vascularizar la parte posterior del mismo. Aunque existen varias arterias
circunferenciales largas, como es el caso de la Arteria Cuadrigeminal (que va
hacia los tubérculos cuadrigéminos), las tres más importantes son las arterias
cerebelosas, que a continuación enumeramos y definimos:
94
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
- Arteria Cerebelosa Póstero-Inferior (PICA): parte de las arterias
vertebrales (aunque en ocasiones lo hace del tronco basilar) y se dirige a
la cara inferior del cerebelo.
- Arteria Cerebelosa Media: suele salir del tronco basilar y se dirige a la
cara anterior del cerebelo.
- Arteria Cerebelosa Superior: nace en el tronco basilar y se dirige a la
cara superior del cerebelo.
POLÍGONO ARTERIAL DE WILLIS
Se trata de un heptágono anastomótico que se establece en la base del cráneo.
Las arterias cerebrales anteriores (ramas terminales de las carótidas internas) van a
dirigirse, por encima de los nervios ópticos, hacia la cisura interhemisférica; y se
anastomosarán entre sí mediante una arteria
de pequeño tamaño que se conoce con el
nombre de arteria comunicante anterior. Por
otra parte otra rama terminal de la carótida,
que es la arteria comunicante posterior, va a
anastomosarse con las arterias cerebrales
posteriores procedentes del tronco basilar.
De esta manera, se establece el Polígono de
Willis, un sistema de protección que evita
que la vascularización cerebral se vea comprometida en determinadas situaciones, pues,
gracias a las anastomosis, un fallo en una arteria puede ser compensado por las otras.
Cabe decir, no obstante, que existen distintos tipos de variaciones del polígono
de Willis. Hay quien dice, incluso, que el polígono de Willis es específico de cada
persona. Lo que sí se conoce con veracidad es el hecho de que esta estructura varía con
la edad y en función de las necesidades de vascularización que exija el encéfalo.
Algunas de las variaciones más frecuentes del polígono de Willis son:
• Comunicante anterior débil: 43%
• Polígono simétrico: 59 %
• Comunicante posterior débil: 25,6 %
• Una sola arteria comunicante posterior débil.
95
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
SISTEMA BASAL
El sistema arterial de la base del cráneo está constituido por una serie de
pequeñas arterias basilares que se introducen en toda la superficie que se corresponde
con la base del cerebro. Existen tres tipos de Arterias Basilares, que son las que
mostramos a continuación:
a) Arterias basilares dependientes de la arteria cerebral posterior.
b) Arterias basilares dependientes de la arteria cerebral media.
c) Arterias basilares dependientes de la arteria cerebral anterior.
A parte de éstas, hemos de decir que las arterias del polígono de Willis también
pueden dar arterias basilares, que se introducirán en el cerebro. Algunas de estas arterias
SISTEMA CORTICAL
La irrigación venosa de la corteza cerebral se va a llevar a cabo gracias a la
acción de tres arterias fundamentales, que son:
a) Arteria cerebral anterior
b) Arteria cerebral media o silviana, que busca la cisura de Silvio.
c) Arteria cerebral posterior, que va hacia el lóbulo occipital.
ARTERIA CEREBRAL ANTERIOR
Es más pequeña que la arteria cerebral media y en cuanto se origina, se dirige
hacia la cisura interhemisférica. Posteriormente rodea el cuerpo calloso. Cuando alcanza
el nivel del esplenio del cuerpo calloso se anastomosa con una rama que procede de la
cerebral posterior. En este caso se establece una anastomosis “boca a boca”.
Ramas de la Arteria Cerebral Anterior
Para poder comprender el territorio que es vascularizado por la arteria cerebral
anterior es necesario especificar las ramas que va a dar a lo largo de su recorrido,
especificando las localizaciones que irrigan cada una de ellas. Estas ramas pueden
agruparse en dos grandes grupos, en función del lugar desde el de donde partan:
96
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
a) Ramas que da la arteria cerebral anterior por su concavidad:
- Arterias estriadas anteriores: son de un tamaño muy reducido y se
dirigen hacia los núcleos grises.
- Ramas para la rodilla y el cuerpo del cuerpo calloso.
- Ramas para los pilares anteriores del trígono.
- Arteria Pericallosa Posterior: se anastomosa con la arteria del esplenio
del cuerpo calloso, rama de la cerebral posterior.
b) Ramas que da la arteria cerebral anterior por su convexidad:
- Arteria calloso-marginal de Foix Hillemond.
- Arteria del lóbulo paracentral: es una rama terminal.
- Arteria estriada media recurrente de Heubner o arteria de la cabeza del
núcleo caudado: Se dirige hacia delante y posteriormente cambia de
dirección y se dirige hacia la zona de la cabeza del núcleo caudado, de
ahí su nombre.
- Arteria orbitaria interna: vasculariza el tercio interno del lóbulo
orbitario.
- Arteria prefrontal.
ARTERIA SILVIANA O CEREBRAL MEDIA
Es la rama más gruesa de la carótida interna y la arteria cerebral de mayor
tamaño. Dado su calibre, cuando se observa en la sala de disección, parece ser la
continuación de la carótida interna mientras que el resto de arterias semejan ramas
colaterales. Como consecuencia de su magnitud, los problemas cerebrales en la
vascularización van a afectar de modo significativo a esta arteria.
La arteria silviana es única, pero en la mayoría de los casos, se divide en dos
ramas: una anterior y una posterior. No obstante en los tratados de anatomía aparece
como una sóla arteria que se ramifica.
Ramas de la Arteria Cerebral Media.
Las ramas que parten de la arteria cerebral media o silviana van a clasificarse en
función de su localización tal y como sigue:
97
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
a) Ramas colaterales superficiales:
- Ramos insulares: van a llevar sangre a la ínsula.
- Arteria orbitofrontal de Foix y Lévy o arteria orbitaria externa: se
encarga de vascularizar los tres tercios externos de la base del lóbulo
orbitario.
- Arteria frontal anterior o de la tercera circunvolución frontal: va a
irrigar el área de Broca.
- Arteria prerrolándica: se introduce en el surco precentral.
- Arteria rolándica: va al surco de Rolando.
- Arteria parietal ascendente parietal anterior o post-rolándica: se dirige
al surco post-rolándico.
- Arteria parietal posterior: va al surco parietal.
- Arteria temporal anterior.
- Arteria temporal media: que es inconstante, y no aparece en todos los
individuos.
- Arteria temporal posterior.
b) Ramas perforantes o arterias centrales: son arterias que parten de la silviana y
que van a perforar la base del cerebro, llegando hasta los núcleos grises:
- Arterias centrales lentículo-estriadas: internas y externas (es importante
la arteria de Charcot). Vascularizan el núcleo caudado, el lenticular y el
tálamo óptico. La mayor parte de las hemorragias intracerebrales se
producen por la arteria de Charcot, ya que es una arteria muy larga que
se puede romper con relativa facilidad al aumentar la presión arterial.
c) Rama terminal:
- Arteria que termina en la circunvolución del pliegue curvo.
ARTERIA CEREBRAL POSTERIOR
Se trata de una arteria que parte del tronco basilar y debe dar la vuelta al tronco
encefálico, de forma que se sitúa entre éste y los hemisferios, más concretamente en la
Hendidura Cerebral de Bichat. Se situará a nivel de la Circunvolución del Hipocampo y
desde ahí se va a dirigir hacia la cisura calcarina, donde dará lugar a la arteria calcarina
(rama terminal). De esta manera, las arterias cerebrales posteriores recorren de adelante
atrás el lóbulo esfenotemporal.
98
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Ramas de la Arteria Cerebral Posterior
Al igual que en casos anteriores, vamos a poder dividir las ramas pocedentes de
la arteria cerebral posterior en función de su localización:
a) Ramas colaterales superficiales:
- Arteria temporal anterior.
- Arteria temporal posterior.
- Arteria temporal media (sólo en ocasiones).
- Arteria del esplenio del cuerpo calloso: se anastomosa con la arteria
pericallosa posterior.
- Arteria del hipocampo o del asta de Ammon: se introduce en la cisura del
hipocampo y vasculariza esta región.
- Ramas circunferenciales que rodean al tronco encéfalo.
- Arterias pedunculares.
- Arteria geniculada.
- Arteria cuadrigeminal.
- Arteria radicular del III par craneal.
b) Arterias de los plexos coroideos: son arterias que constituyen los plexos
coroideos mediante anastomosis. De las arterias cerebrales posteriores proceden:
- Arteria coroidea posterior.
- Arteria coroidea media.
c) Región profunda (formada por tálamo e hipotálamo):
- Arterias interpedunculares.
- Arterias tálamo-tuberianas.
- Arterias tálamo-geniculadas.
d) Ramas terminales:
- Arteria Calcarina.
SISTEMA VENTRICULAR O COROIDEO
Existen tres arterias, que ya conocemos, que forman el sistema ventricular o
coroideo. Estas arterias son la coroidea anterior (rama de la carótida interna), la
coroidea media y la coroidea posterior (estas dos últimas ramas colaterales de la arteria
cerebral posterior).
99
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Hay tres aspectos que hay que tener en cuenta en lo que se refiere al sistema
ventricular o coroideo:
• En primer lugar, hay que dejar de manifiesto el hecho de que las tres arterias se
anastomosan para dar lugar a los plexos coroideos.
• En segundo lugar, cabe decir que la arteria coroidea anterior es la más
importante de las tres. No obstante, presenta un tamaño variable, hasta el punto
de que, en ciertos individuos en los que las arterias coroideas posterior y media
son especialmente pequeñas, la arteria coroidea anterior puede llegar incluso
hasta el agujero de Monro.
• Finalmente, es preciso señalar la existencia de unas ramas perforantes que parten
de la arteria coroidea anterior cuya función va a ser la vascularización de la
cápsula interna (El Brazo Conjuntival posterior, que separa el núcleo lenticular
del tálamo). Es esta una zona por la que discurren numerosas fibras relacionadas
con el movimiento voluntario, de ahí que obstrucciones de esta arteria puedan
dar lugar a parálisis de mayor o menor gravedad en función del tipo de
obstrucción.
TERRITORIOS DE VASCULARIZACIÓN DE LAS ARTERIAS CEREBRALES
Es tremendamente importante para la práctica clínica conocer los distintos
territorios que van a estar irrigados por distintas arterias. Esta importancia radica en el
hecho de que obstrucciones en distintas arterias provocarán atrofias en distintos lóbulos
cerebrales, causando una serie de síntomas específicos que el médico debe reconocer.
Así, procederemos a establecer los distintos territorios de cuya vascularización se
encargan las distintas arterias cerebrales:
• Arteria Cerebral Anterior: vasculariza la cara interna el lóbulo frontal, así como
el tercio interno del lóbulo orbitario. También irriga el borde superior de la cara
externa de los hemisferios cerebrales.
• Arteria Silviana: irriga la mayor parte de la cara externa de los hemisferios
cerebrales, pero dejando el borde superior (que pertenece a la arteria cerebral
anterior) y el inferior (irrigado por la cerebral posterior).
• Arteria Cerebral Posterior: lleva sangre al lóbulo occipital y a la región
esfenotemporal.
100
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
SISTEMA DE LOS NÚCLEOS GRISES
Existen una serie de arterias que vascularizarán los núcleos grises que se sitúan
en el interior del cerebro. No obstante, debemos especificar que se trata de arterias que
constituyen ramas colaterales o terminales de otras que se encargan también de la
vascularización de la corteza cerebral. Por ello, estudiaremos la vascularización de cada
núcleo por separado:
1. Tálamo óptico: las arterias que se encargan de la vascularización del tálamo óptico
de cada uno de los hemisferios son:
- Arteria Cerebral Posterior.
- Arteria Comunicante Posterior.
- Arteria Cerebral Media.
- Arteria Coroidea Anterior.
2. Núcleo estriado: el estriado va a estar compuesto por tres unidades diferentes, que
serán vascularizadas por arterias distintas. Así tenemos:
a) Cabeza Del núcleo caudado: vascularizada por las arterias que a
continuación se presentan:
- Arteria Cerebral Anterior
- Arteria Recurrente de Heubner.
b) Cuerpo del núcleo caudado, putamen y palidum externo: que van a estar
irrigados por una única arteria, que es:
- Arteria Cerebral Media.
c) Palidum interno: la única vascularización que recibirá va a provenir de
una sóla arteria:
- Arteria Coroidea Anterior.
3. Cápsula Interna: va a ser vascularizado por una única arteria, que se presenta a
continuación:
- Arteria Cerebral Media: que lleva la sangre hasta el brazo conjuntival
anterior
101
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
V
A
S
C
U
L
A
R
I
Z
A A
C R
I T
Ó E
N R
I
A
L
102
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VASCULARIZACIÓN VENOSA DEL ENCÉFALO
Al igual que el encéfalo presenta una serie de arterias características que llevan
la sangre a las distintas regiones del mismo, va a presentar también una serie de venas
para drenar la susodicha sangre. Ahora procederemos a su estudio.
En relación a la clínica, debemos decir que, a pesar de que las patologías más
frecuentes o diagnosticados son los problemas arteriales, existen autores que consideran
que dichos problemas van a ser el resultado de otros previos en el sistema venoso.
103
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CARACTERÍSTICAS GENERALES DE LAS VENAS CEREBRALES
Del mismo modo que ocurre con las arterias, y dadas sus características
especiales, el encéfalo va a poseer una serie de venas con unas particularidades
específicas. Las más importantes de dichas particularidades son las que enumeramos a
continuación.
• Ofrecen pocas analogías con el sistema arterial.
• No siguen un surco, saltan sobre varios y suelen mantenerse en superficie.
• Cuando arteria y vena van juntas, la arteria siempre se sitúa en el plano
profundo.
• La pared venosa es muy fina.
• Sólo las venas más gruesas presentan fibras musculares.
• Las venas cerebrales carecen de válvulas, pues el flujo sanguíneo va a
depender de la posición de la cabeza, por lo que dicho no será
unidireccional.
• La red venosa superficial está ampliamente anastomosada.
• Las venas intraparenquimatosas profundas son terminales.
CLASIFICACIÓN DE LAS VENAS
Para clasificar las venas vamos a centrarnos en su localización en relación con el
conjunto del encéfalo. De esta manera tendremos:
• Venas superficiales o corticales.
• Venas centrales: recogen sangre de los plexos coroideos y ventrículos.
• Sistema venoso basilar: situado en la base del cerebro.
Todos los sistemas venosos presentan una característica común, que es el hecho
de que van a drenar a los senos venosos. A su vez, la corriente más importante de los
senos venosos va a dirigirse hacia el seno lateral o sigmoideo, y desde él, a la vena
yugular interna. Se trata, pues, de un sistema triple que termina en la yugular.
Existen pequeñas anastomosis extra-intracraneales (entre circulación venosa del
interior y del exterior del cráneo), que presentan una cierta importancia, pues pueden
dar paso a bacterias u otros elementos entre el endocráneo y el exterior del mismo.
104
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Cabe señalar, no obstante, que además de estas vías, que son las más comunes,
van a aparecer otras de menor relevancia, como es el caso de la que incluye a la vena
angular, la vena oftálmica y el seno cavernoso.
VENAS EMISARIAS
Son bastante variables, y reciben este nombre porque comunican la circulación
de un seno venoso con partes extracraneales. Normalmente son muy pequeñas y suelen
estar situadas en las cercanías del seno longitudinal superior y del seno lateral. Su
importancia y calibre dependen del individuo de estudio, pero por norma general, las
más frecuentes son las parietales y las de la escama del temporal.
VENAS SUPERFICIALES O CORTICALES
Las venas superficiales son también bastante variables y se nombran en relación
topográfica con el lóbulo o cisura en la que se localiza. Así, en la cisura de Silvio, nos
encontraremos la vena silviana, la vena prerrolándica en la cisura prerrolándica,…. Si la
vena “salta” sobre varias circunvoluciones, se le da el nombre del lóbulo al que
pertenece.
La forma de drenar de las venas en los senos es distinto en la zona frontal que en
la occipital. En el lóbulo frontal, las venas terminan a favor de la corriente, mientras que
a nivel occipital las venas drenan contracorriente, una característica que es el resultado
del desarrollo del cerebro.
Como sabemos, por norma general, las venas superficiales están anastomosadas,
pero existen unas anastomosis mucho más constantes que otras, de forma que algunas
de ellos adquireron relevancia quirúrgica como punto de localización topográfica. Las
anastomosis venosas más frecuentes que nos podemos encontrar son:
• Vena anastomótica de Labbe: anastomosis entre la vena silviana superficial y el
seno lateral.
• Vena anastomótica de Trolard: une el seno venoso longitudinal superior con la
vena silviana
• Canal colateral parietal: vena que parte del seno longitudinal superior y termina
en la vena anastomótica de Labbe.
105
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VENAS CENTRALES
A nivel del agujero de Monro existe una formación venosa muy importante, pues
en esta localización se unen, por un lado, la vena del Septem lúcidum, por otro la opto-
estriada y finalmente, por otro, la vena coroidea. Una vez que estas tres venas se han
unido (por detrás del agujero de Monro), se forma un colector que se dirigirá hacia atrás
y dará lugar a la Vena Cerebral Interna, que terminará, a su vez, en la ampolla o gran
vena de Galeno. De esta manera, y como es lógico, existen dos venas cerebrales
internas, una en cada hemisferio.
SISTEMA BASILAR
A la base del cráneo van a llegar una serie de venas que se van a anastomosar
dando lugar a lo que se conoce con el nombre de Polígono de Trolard. Por una parte,
llegan las venas cerebrales anteriores. La vena silviana, que viene por la cisura de Silvio
se unirá con las cerebrales anteriores,
dando lugar a un gran confluente
venoso, que se dirige hacia atrás y da
lugar a la vena basilar de Rosental.
Las venas cerebrales anteriores se
anastomosan entre sí gracias a la
vena comunicante anterior. Las
basilares se anastomosarán también mediante la vena comunicante posterior. Con todas
estas estructuras se configura por completo el polígono de Trolard del que ya hemos
hablado.
106
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
V
A
S
C
U
L
A
R
I
Z
A
C
I
Ó
N
V
E
N
O
S
A
107
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Una vez estudiados todos los elementos que componen el cerebro, es hora de
estudiarlos en su conjunto. Para ello vamos a realizar una serie de cortes en el mismo,
que serán de dos tipos:
• Cortes de Charcot: se trata de cortes frontales o coronales.
• Cortes de Fleshing: son cortes horizontales.
Dado que cada corte va a ser distinto, se darán unas características generales que
permitan identificar la región. Por ello debemos localizar:
a) El cuerpo calloso. Debemos fijarnos también en la presencia o no del Septum
lúcidum y el trígono. Como ya sabemos en la parte anterior el Septum es de
mayores dimensiones y el trígono relativamente pequeño, situación que se
invierte a medida que nos dirigimos a la zona posterior.
b) Ventrículos laterales.
c) Núcleos grises.
d) En los cortes de Charcot debemos identificar la forma de los lóbulos frontal y
occipital. La base del polo frontal es más o menos plana y los lóbulos están
juntos. Los lóbulos occipitales, por el contrario, están más separados y rematan
de forma oval, dado que el cerebelo se introduce a modo de cuña entre ambos
lóbulos.
ACLARACIONES SOBRE LOS CORTES DE CEREBRO
En los cortes de cerebro nos vamos a encontrar multitud de estructuras
conocidas, pero será necesario realizar algunas aclaraciones acerca de ellas. Del mismo
modo, aparecerán nuevas estructuras que nombraremos y explicaremos:
• Existe una lámina de sustancia gris entre el putamen y la ínsula que se conoce
con el nombre de Claustrum o Antemuro.
• Podemos diferenciar una estructura de sustancia gris próxima al tálamo que se
conoce con el nombre de Sustancia Enrejada o Aparrillada de Arnold (forma una
red al lado de los tálamos).
108
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• A cada lado del cuerpo calloso quedan unas superficies de sustancia blanca a
cada lado que se conocen con el nombre de Centros Semiovales, y que, juntos,
forman el Centro Oval del Cuerpo Calloso.
• El Septem lúcidum está formado, en realidad, por dos tabiques que se juntan, por
lo que en ciertas localizaciones puede observarse una pequeña cavidad originada
por estos dos tabiques.
• Existe una región de sustancia blanca situada entre el núcleo lenticular y el
conjunto núcleo caudado/tálamo. Es una zona por la que bajan las fibras motoras
voluntarias y recibe el nombre de Cápsula Interna. Existen dos cápsulas más,
que son la Cápsula Externa (entre el núcleo lenticular y el claustrum) y la
Cápsula Extrema (entre el claustrum y el lóbulo de Ínsula).
• Por debajo del núcleo lenticular y encima de la cola del núcleo caudado aparecen
fibras que pertenecen a la cápsula interna.
• Otras estructuras a destacar son las Fibras de Asociación. Se trata de estructuras
que ponen en contacto distintas partes del cerebro, y que se denominan en
función de su localización topográfica.
• Algo a destacar es el hecho de que el núcleo caudado y el lenticular se
encuentran separados por la cápsula interna, pero existen entre ellos una serie de
fibras que los ponen en contacto. Estos regueros de fibras forman los Puntos
Putámino-Caudados.
• A nivel de la cisura calcarina nos encontramos con una lámina de color blanco.
En esta zona aparece, no obstante, una estría que se conoce con el nombre de
Estría de Genaris, por lo que la corteza cerebral en las proximidades de la cisura
calcarina se denomina Corteza Estriada. En esta región se sitúa el área primaria
de la visión.
• Entre los tubérculos cuadrigéminos queda un espacio en el que se va a asentar la
glándula epífisis. De ahí, que sea conocido con el nombre de Lecho de la
Epífisis.
109
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
110
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Una vez estudiadas las regiones de la médula espinal y conocida su morfología,
debemos profundizar en los aspectos referentes a su funcionalidad. De esta manera,
podemos observar la médula desde dos puntos de vista diferentes:
En primer lugar, cabe decir que la médula espinal es una zona de paso, por lo
que por ella van a pasar todas las fibras que comunican encéfalo y médula. Todas las
órdenes que el SNC envía al SNP van a discurrir por la médula. De igual forma, toda la
información sensitiva que recibimos periféricamente pasará por la médula para llegar
hasta el encéfalo. Así, en la médula nos vamos a encontrar con elementos ascendentes y
descendentes. Los sistemas ascendentes llevan información al encéfalo, por lo que son
de naturaleza sensitiva (son aferentes), mientras que las fibras descendentes son
fundamentalmente motoras, eferentes.
En segundo lugar, es de destacar que la médula presenta sus propias neuronas,
específicamente situadas en la sustancia gris. Esas neuronas van a ser elementos que por
un lado inervan de forma motora una gran parte del cuerpo y las vísceras
(motoneuronas) y que por otro recogen la sensibilidad del cuerpo y las vísceras
(neuronas sensitivas). Dichas neuronas presentan una serie de fibras que las conectan
unas con otras, de forma que se asocian las distintas partes de la médula espinal. Estas
estructuras reciben el nombre de Fibras de Asociación. Los núcleos neuronales pueden
ser, pues, motores, sensitivos y vegetativos.
Gracias a estas características, podemos razonar el hecho de que en los lugares
donde hay mayor número de células, exista más sustancia gris, tal y como ocurre en los
engrosamientos cervical y lumbar. En cuanto a la sustancia blanca, podemos decir que
es más abundante cuanto más craneal sea el corte. Esto se debe a que por la región
cervical pasan todas las fibras que llegarán a toda la médula. Así, en los cortes, el
número de fibras irá disminuyendo a medida que éstas hagan sinapsis en sus
correspondientes destinos. Gracias a estas características, podemos localizar la región en
la que se realiza un corte de la médula, por lo menos de forma aproximada.
SUSTANCIA GRIS DE LA MÉDULA ESPINAL
Para comprender la distribución de la sustancia gris en la médula espinal
diremos que, como norma general, las neuronas que se sitúan en el asta anterior tienen
una función principalmente motora, las de la zona media funciones vegetativas y las que
se sitúan en las astas posteriores, sensitivas.
111
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
ASTAS ANTERIORES
En las astas anteriores de la médula espinal nos encontramos con
motoneuronas, acompañadas por interneuronas (que no son ni sensitivas ni motoras).
Existen dos grandes tipos de motoneuronas, cuya existencia se debe a las diferencias
entre las dos clases principales de músculos estriados:
a) Motoneuronas Alfa (Mα): se encargan de inervar la musculatura de
trabajo. Dentro de la musculatura de trabajo, podemos distinguir también
dos grandes grupos, por lo que van a existir también dos tipos de
motoneuronas alfa:
- Motoneuronas alfa para músculos fásicos o rápidos: los
músculos fásicos están compuestos por fibras musculares pálidas,
que se contraen rápidamente, pero que se agotan enseguida a causa
de su metabolismo glucolítico.
- Motoneuronas alfa para músculos tónicos o lentos: son más
resistentes que los fásicos, gracias al empleo de un metabolismo
oxidativo.
Algo a destacar en relación con las motoneuronas alfa, es que podemos
variar la proporción de motoneuronas alfa fásicas y tónicas con el
ejercicio, por lo que podemos decir que dichas proporciones se adaptan a
los músculos. Esto se acoge perfectamente a la sentencia clásica que dice
que “La función hace al órgano”.
b) Motoneuronas Gamma (Mγ) : los músculos que inerva este tipo de
neuronas se contraen, pero no ejercen trabajo físico, sino que regulan un
receptor que es el huso neuromuscular, cuya función es determinar el
grado de contracción de los músculos. De ahí que esta musculatura se
denomine intrafusal.. Hoy en día también se les aplica la división en
fásicas y tónicas.
Todas estas motoneuronas se sitúan en el asta anterior de una forma ordenada,
formando dos grandes grupos de columnas que se conocen con el nombre de columnas
motrices. Algunas se sitúan en la parte interna del asta y otras en la parte externa, por lo
que nos encontraremos con unas columnas mediales y otras laterales. Las columnas
mediales van a inervar los músculos del cuello y del tronco, mientras que las laterales
inervarán la musculatura de las extremidades superiores e inferiores. Las motoneuronas
112
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
de las columnas laterales se encontrarán sólo en los engrosamientos, mientras que las
correspondientes a las columnas mediales serán constantes a lo largo de toda la médula.
Las neuronas de los músculos flexores se sitúan profundamente en el asta anterior,
mientras que las de los extensores se sitúan más superficialmente.
Existen dos Columnas Mediales, que son las que enumeraremos a continuación,
explicando las partes del cuerpo a las que dan inervación:
• Columna Ántero-medial: se corresponde con las motoneuronas extensoras
de tronco y cuello.
• Columna Póstero-medial, con motoneuronas flexoras de los músculos
ántero-laterales de tronco y cuello.
En cuanto a las Columnas Laterales, cabe decir que existen cuatro de ellas, que
son las que a continuación presentamos:
• Columna Anterior: encargada de la inervación de hombro y pelvis.
• Columna Lateral: que inerva el brazo en el miembro superior y el muslo en
el miembro inferior.
• Columna Póstero-lateral: que dará inervación al antebrazo en el miembro
superior y a la pierna en el inferior.
• Columna Retropóstero-lateral: da inervación a manos y pies.
Existen dos grupos más de motoneuronas que forman unos núcleos en la médula
espinal. Uno de ellos se sitúa en el centro de las columnas y se conoce con el nombre de
Núcleo Central, mientras que el otro se encuentra en la parte más anterior y lateral del
asta anterior, y se conoce como Prolongación Lateral del Asta Anterior.
El núcleo central sólo se encuentra en C4 y en alguna parte de C5, y es el más
importante de los núcleos de motoneuronas de la médula, pues inerva al diafragma. De
él sale el nervio frénico, de ahí que el núcleo se conozca también con el nombre de
Núcleo Frénico.
La prolongación lateral del asta anterior presenta, a su vez, las motoneuronas
responsables de la inervación del trapecio y del esternocleidomastoideo. Son, por tanto,
neuronas correspondientes al nervio espinal, y se encuentra en los primeros segmentos
cervicales, específicamente en C1 y C2. No obstante, debemos decir que, dado que el
nervio espinal (XI par craneal) presenta dos grandes funciones, va a tener dos orígenes
distintos. Por un lado es el nervio motor de los músculos de la laringe y permite la
función del habla, y por otro, es el nervio que mueve la cabeza (nervio cefalogiro) ya
113
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
que inerva al trapecio y al esternocleidomastoideo. De esta manera, presenta un origen
en núcleos bulbares para la inervación de los músculos de la laringe y otro en la
prolongación lateral del asta anterior para la rama que inerva el trapecio y el
esternocleidomastoideo.
En el asta anterior, por cada motoneurona, podemos encontrar unas quince o más
interneuronas, de las cuales algunas presentan una mayor importancia clínica, como es
el caso de las neuronas inhibidoras de Rensu, que se encargan de inhibir a una
motoneurona estimulada y en funcionamiento (dicha motoneurona manda una rama
colateral a la neurona inhibidora)
ASTAS POSTERIORES
Las astas posteriores de la médula espinal desarrollan funciones sensitivas, y las
neuronas que las llevan a cabo se van a organizar en una serie de núcleos.
De esta manera, en la región más posterior del asta posterior de la médula nos
encontramos la Zona Marginal de Lissauer, que es una localización medular
tremendamente rica en fibras y muy poco en células; hasta tal punto que algunos autores
consideran que dicha zona marginal es una región compuesta por fibras, mientras que el
lugar donde se situarían las células se denominaría núcleo póstero-marginal.
Por delante de la zona marginal de Lissauer podemos observar otra estructura,
que se conoce con el nombre de Núcleo Gelatinoso de Rolando, y que está relacionada
con las vías del dolor.
En una parte más anterior, se puede distinguir el Núcleo Sensitivo Propio o
Principal del Asta Posterior, en el que se localizan todas las neuronas de la sensibilidad
salvo las que se encargan de transmitir la información de dolor.
Finalmente, por delante del núcleo sensitivo nos encontraremos dos nuevos
núcleos de considerable tamaño que son:
• Núcleo de Clarke (núcleo interno).
• Núcleo de Betcherew (núcleo externo).
Estos dos núcleos se sitúan en la base del asta posterior, y se relacionan con
funciones propioceptivas (aquellas funciones sensitivas que permiten conocer lo que
ocurre, por ejemplo, en nuestro aparato locomotor: si los músculos están contraídos o
relajados, en contracción o relajación, situación de palancas óseas,….)
114
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La clasificación por núcleos que acabamos de explicar es tremendamente útil
para la práctica clínica. No obstante, para la investigación anatómica y fisiológica
actual, se tienen en cuenta también las fibras que llegan y parten de dichos núcleos, por
lo que aparece una nueva división de la médula en áreas que se conoce como
Laminación Medular de Rexed, de la cual hablaremos más adelante.
ZONA INTERMEDIA DE LA MÉDULA ESPINAL
La región intermedia de la médula espinal debe ser asociada, desde el punto de
vista médico, a funciones vegetativas. En esta zona vamos a poder distinguir dos
grandes columnas:
• Columna intermedio-medial: aparece a lo largo de toda la médula espinal e
inerva a todos aquellos órganos que poseen inervación simpática y que
están extendidos por todo el cuerpo. Un ejemplo claro serían las glándulas
sudoríparas o el músculo horripilador (que pone los pelos de punta o la
carne de gallina). La parte anterior de dicho núcleo es motora y la posterior
sensitiva.
• Columna intermedio-lateral: Se encuentra fundamentalmente a nivel de la
región dorsal y va a dar inervación a una gran cantidad de órganos
vegetativos, de forma que existen unos determinados centros en la médula
que se corresponden con distintas regiones del organismo:
- Centro Cilioespinal o de Budge (C8, T1 y T2)
- Centro bronco pulmonar (T3, T4 y T5)
- Centros esplácnicos abdominales (desde C6 hasta C11): inervan
simpáticamente las vísceras abdominales (centro simpático
medular, junto con los esplácnicos pelvianos)
- Centros esplácnicos pelvianos (T12, L1 y L2): va a inervar los
intestinos de forma simpática.
- Centros pelviperineales (desde S1 hasta S4): también inerva
vísceras pélvicas. Es la única zona de inervación parasimpática en
la médula espinal. No obstante, cabe decir que las vísceras no
pélvicas van a tener también inervación parasimpática, pero en
este caso, la inervación le llega del neumogástrico. De esta forma,
observamos que los centros simpáticos se sitúan todos en la
115
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
médula espinal, mientras que los parasimpáticos se sitúan en la
médula, pero también el tronco del encéfalo.
Funcionamiento básico de las vías simpáticas y parasimpáticos.
Los nervios simpáticos son sensitivos y motores. De esta forma, cuando en un
determinado órgano se recoge la información, ésta se va a dirigir hacia un ganglio
vegetativo. Cuando la información llega a la médula, las neuronas motoras simpáticas
que parten de ésta, pueden “atravesar” la cadena simpática para dirigirse al ganglio
correspondiente en el que se sitúa la segunda neurona de la vía simpática.
Las vías parasimpáticos son similares, pero presenta una diferencia: las segundas
neuronas motoras se sitúan en ganglios parasimpáticos próximos al órgano, o incluso
incluidos en sus propias estructuras (como ocurre con algunos ganglios uterinos, por
ejemplo). En el sistema parasimpático pélvico, las neuronas no se dirigen hacia el asta
posterior de la médula, sino que penetran en la misma por el asta anterior y luego se
dirigen a la zona posterior. Se trata de un caso especial que se salta todo tipo de reglas y
cuya razón es totalmente desconocida.
116
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
ORGANIZACIÓN LAMINAR DE REXED
Una vez conocida la citoarquitectura de la sustancia gris y las funciones que
desarrollan cada una de las regiones que la componen, podemos comprender de mejor
modo la organización de Rexed. Fue éste un científico que desarrolló un sistema de
división en áreas de la médula teniendo en cuenta no sólo como estaban distribuidas las
neuronas sino también donde sinaptaban. Así se creó la Laminación de Rexed, que
divide la médula en diez láminas., de forma que la lámina I comienza en el área
posterior, y la X acaba en la zona central (región del conducto ependimario). Estudiando
las láminas en relación con cada una de las regiones que conocemos tenemos:
• En el asta posterior: existen seis láminas (concretamente de la I a la VI). El
conjunto de las columnas que conocemos de esta región conforman la
Lámina IX (que representa, por tanto, el conjunto de todas las
motoneuronas).
• En la Zona Intermedia y el Asta Anterior: en ellas se sitúan las láminas VII y
VIII. Se trata de láminas compuestas, en su inmensa mayoría, por
interneuronas (que no son ni sensitivas ni motoras)
• Zona Ependimaria: alrededor del epéndimo se sitúa la lámina X.
La referencia de Rexed es la empleada en los trabajos de investigación actuales
que se refieren a la médula espinal, por lo que su conocimiento es de especial interés
para poder comprender las publicaciones científicas.
IMPORTANCIA CLÍNICA DE LA CITOARQUITECTURA DE LA
SUSTANCIA GRIS MEDULAR
Conocer La citoarquitectura de la sustancia gris medular es de especial
relevancia para el médico porque casi todos los músculos están inervados por varias
raices. Con las tablas de músculos-inervación, a partir de los músculos afectados,
podemos saber que región de la médula está afectada gracias a métodos de exploración
clínica.
De esta manera, podremos identificar lesiones medulares porque, dado que
existen segmentos que invervan varios músculos, si está afectado un sólo músculo, será
defecto de un tronco; mientras que si están afectados todos los músculos inervados por
una raíz medular, se podrá concluir que nos encontramos ante una lesión medular.
117
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
LA SENSIBILIDAD
En lo que se refiere a la sensibilidad, hemos de decir que el cuerpo está dividido
en segmentos, que se denominan Dermatomas. Así, se define un dermatoma como un
segmento de piel inervado por una determinada raíz raquídea. Por ello, para llevar a
cabo una correcta exploración de la sensibilidad debemos comprobar todos los
dermatomas, pues es posible la existencia de un dermatoma alterado mientras que los
demás permanecen normales. El concepto de dermatoma implica también el hecho de
que cada zona de la piel está inervada por un par raquídeo diferente, de forma que
podemos llegar a un diagnóstico.
VÍAS DESCENDENTES DE LA MÉDULA ESPINAL
La mayor parte de las vías descendentes de la médula son vías motoras que van a
enviar órdenes a los órganos efectores. En la práctica clínica, estas vías motoras se
valorarán en una escala del 0-5, que mide la fuerza muscular en función del nivel de
contracción de los músculos. Los distintos valores y su significado los expresamos a
continuación:
• Valor 0: Contracción nula.
• Valor 1: Existe contracción, pero es inefectiva, dado que, aunque el músculo
se contrae, no es capaz de mover el segmento.
• Valor 2: en el nivel dos existe contracción, pero insuficiente para vencer la
gravedad.
• Valor 3: en este caso, la contracción muscular es capaz de vencer la
gravedad.
• Valor 4: en el cuarto nivel, puede vencer la gravedad, pero la fuerza normal
se ve disminuida.
• Valor 5: nivel de contracción y fuerza muscular normal.
La que se detalla, es la valoración internacional, y cuando exploramos a un
individuo testamos, mediante este sistema, la contracción de los principales músculos.
118
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VÍAS MOTORAS
Las vías descendentes motoras de la médula espinal van a poder clasifican en
dos grandes grupos, que a continuación exponemos:
• Vías de la motilidad voluntaria: se conocen clásicamente con el nombre de
vías piramidales y vías geniculadas.
• Vías del movimiento asociado al movimiento voluntario: que también
podrían llamarse (aunque no sean estrictamente lo mismo), vías
involuntarias. Estas vías se conocen clásicamente con el nombre de vías
extrapiramidales. Cabe decir, en lo referente a la denominación de “vías
involuntarias”, que hoy día sabemos que no existe ningún movimiento
voluntario puro, ni tampoco un movimiento involuntario totalmente puro.
Esto se debe a que, cada vez que se realiza un movimiento voluntario, se
mueven muchos músculos, pues cambia el centro de gravedad del cuerpo, y
éste necesita adaptarse a los cambios de posición. Además, cualquier
movimiento voluntario puede llegar a ser automatizado.
VÍAS DEL MOVIMIENTO VOLUNTARIO
Desde el punto de vista del movimiento voluntario las vías se clasifican en
piramidales y geniculadas. Las vías geniculadas no llegan a la médula, sino que se
quedan en el encéfalo. Esto se debe a que este tipo de vías se encargan del movimiento
voluntario de los músculos inervados por los pares craneales, cuyas raíces se sitúan a
nivel encéfalico. Las vías piramidales, por el contrario, se van a encargar del
movimiento voluntario de los músculos inervados por los pares raquídeos.
VÍA PIRAMIDAL
Las vías piramidales fueron clasificadas, históricamente, en dos tipos diferentes,
tal y como a continuación mostramos:
• Vía piramidal directa.
• Vía piramidal cruzada (se sitúa en el cordón lateral de la médula espinal).
119
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Ambas primeras de las vías son cruzadas y representan, al menos, el 99 % de la
vía piramidal. Como posteriormente aparecieron unas fibras que no se cruzaban, a las
que se le dio el nombre de Vía Piramidal Homolateral. No obstante, estas fibras se
corresponden con menos del 1 % de las que componen las vías piramidales, por lo que,
aunque su conocimiento puede interesar desde el punto de vista teórico, su existencia no
es significativa en casos prácticos.
La diferencia entre las vías piramidales directa y cruzada reside en el lugar de
cruce, así como en el número de fibras. La cruzada supone el 90% de las fibras de las
vías piramidales, mientras que la únicamente se corresponde con el 9% de las
susodichas fibras.
Para poder comprender el funcionamiento de estas vías, hemos de decir que se
considera que la realización de un movimiento voluntario consta de tres niveles:
1. Nivel superior o de Estrategia(objetivo del movimiento): de esta función se
encarga, según se cree, la zona prefrontal del lóbulo frontal. Este área es la
región de la motivación.
2. Nivel medio o de Táctica: Se trata de un sistema que se encarga de la
planificación y las secuencias espacio-temporales necesarias para la realización
del movimiento. Planifica, pues, aquellas neuronas que deben activarse.
3. Nivel inferior o de Ejecución: supone la activación de las neuronas generadoras
del movimiento. Se trata de la vía motora en sí. El sistema ejecutivo general
consta de dos neuronas. Existe una primera, que es la neurona motora superior, y
una segunda que se conoce como neurona motora inferior. A nivel médico,
sabemos que cuando existe una parálisis puede ser producida en una de estas dos
neuronas, y será necesario localizar tanto la neurona afectada, como la región de
la misma en la que se produce la lesión.
El Nivel Inferior o de Ejecución.
La primera neurona se sitúa en la corteza cerebral, en el área 4 (circunvolución
frontal ascendente), por lo que algunos autores la conocen como neurona motora
cortical. De esta manera, en esta región, vamos a poder distinguir diferentes áreas del
cerebro que presentan las neuronas que darán inervación a las diferentes partes de
cuerpo. Dado que ciertas localizaciones corporales precisan mayor inervación que otras,
en una representación gráfica de la circunvolución frontal ascendente en relación con las
120
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
áreas corporales a las que inerva, podemos observar un “hombre” acostado y
desproporcionado en cuanto a sus miembros que se conoce como el Homúnculo de
Penfield. El área cuatro es también la responsable de los movimientos más finos,
función desempeñada por unas células que existen en esta región y que se conocen con
el nombre de Células Piramidales de Betz, que son las más grandes del organismo.
La segunda neurona es una motoneurona de la médula, con la cual sinapta la primera
neurona. Ésta, tiene dos formas de llegar hasta la médula: puede cruzarse en la
decusación del bulbo y dirigirse directamente hacia la segunda neurona para sinaptar
con ella (que enviará el impulso a un músculo estriado), dando lugar a la Vía Piramidal
Cruzada; o bien cruzándose en cada segmento medular, sin hacerlo en la decusación
de Misticheli, dando lugar a la Vía Piramidal Directa. Como podemos observar, a
causa de estos cruzamientos neuronales (decusaciones), un hemisferio se encarga del
control motor del lado contrario del cuerpo.
121
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La primera neurona pasa por el brazo conjuntival posterior de una forma
ordenada. A nivel mesencefálico, la vía
piramidal se sitúa en el pie, por delante de
la sustancia negra (más concretamente en
la zona media). Por la parte externa se
sitúan los Fascículos Témporo-Pontinos, y
por la interna la Vía Geniculada.
Posteriormente, la vía piramidal desciende
por los rodetes de la protuberancia y las
pirámides bulbares, para llegar finalmente
a la médula.
En lo que se refiere a la segunda neurona, denominada por Sherrington como
Vía Final Común, cabe decir que cualquier músculo que se contrae voluntariamente,
precisa de dicha neurona. Es la neurona motora inferior, que establece la conexión entre
la primera neurona y el músculo efector.
Aplicaciones clínicas del conocimiento de las vías descendentes motoras.
Como médicos debemos saber que, cuando existen parálisis musculares
voluntarias, van a ser el resultado de lesiones de la neurona motora superior o inferior
de la vía piramidal. Por ello, en primer términos debemos reconocer cuál es la neurona
afectada, y posterioremente, hemos de localizar dicha lesión.
1. Reconocimiento de la neurona afectada: para reconocer las afecciones de
las motoneuronas hemos de decir que los síntomas que producen son la
paresia (parálisis parcial) o la parálisis total. No obstante, no todas las
parálisis son iguales, sino que existen dos tipos:
a) Parálisis Flácida: además de no poder realizar el movimiento
voluntario del músculo, éste se encuentra flácido. Es
característica de las lesiones de neurona inferior, pues las
neuronas del asta anterior de la médula son las encargadas del
trofismo muscular.
b) Parálisis Espástica: los músculos se encuentran en estado de
contracción, aunque el paciente no pueda moverlos de forma
voluntaria. Esta característica de los músculos se denomina
122
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
espasticidad, y es proporcional a la extensión (si le extendemos el
músculo al paciente, la contracción aumentará). Se trata de una
parálisis característica de primera neurona.
Además de lo anteriormente enunciado, cabe decir que en las lesiones de
primera neurona aparecen una serie de reflejos que, por norma general,
son patológicos. El más conocido es el Reflejo de Babinski, que
consiste en estimular la planta del pie. El paciente da positivo al reflejo
de Babinski cuando responde al estimulo de la planta del pie con una
extensión en abanico de los dedos (la respuesta normal sería la flexión de
los mismos). Es de destacar, sin embargo, que en los recién nacidos, y
hasta el año de edad, el reflejo de Babinski es positivo; dado que la vía
piramidal es la última en aparecer y la última en la que se produce la
mielinización. La espasticidad y el reflejo de Babinski se producen
porque, acompañando a la piramidal, existe otra vía que recibió el
nombre de Vía Parapiramidal. Hoy sabemos que es la Vía Córtico-
Espinal, que discurre entre las fibras de la piramidal, de forma cuando se
lesiona la primera neurona, aparece parálisis y fenómenos de liberación
de sustancias que antes permanecían ocultas por fibras. Para concluir,
podemos citar otro ejemplo patológico, como es el caso del Reflejo de
triple retirada, que es normal cuando existe algún estímulo dañino, pero
que en afecciones de motoneuronas actúa con sólo tocar la piel.
2. Reconocimiento del área de neurona lesionada: debido a la complejidad
para situar la región afectada de la motoneurona no funcional, este
apartado se explicará un poco más adelante en nuestro estudio.
VÍA GENICULADA
La vía geniculada también se denomina córtico-nuclear y es una vía descendente
cruzada toda ella, que presenta únicamente presenta una excepción (que no vamos a
estudiar)
Para poder comprender la vía geniculada, es preciso situar los núcleos que
suponen los orígenes reales de los pares craneales, dado que dicha vía es la que se
encarga de los movimientos voluntarios producidos por ellos. Así tenemos que:
123
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• El núcleo del Trigémino (V par), se sitúa a nivel de la protuberancia o puente de
Varolio.
• En el bulbo se encuentra el núcleo motor del Hipogloso (XII par)
• En el mesencéfalo, podemos situar los núcleos de los pares craneales III y IV.
• El núcleo del VII par se localiza en el Surco Bulbopóntico.
El esquema base de la vía geniculada va a estar compuesto también por dos
neuronas: una primera, superior o cortical, y una segunda que pertenece al tronco
encefálico (neurona motora inferior). La
neurona cortical o neurona motora
superior pasa por la rodilla de la cápsula
interna (de ahí el nombre de geniculada).
Posteriormente, pasará por la parte
interna de los pedúnculos cerebrales, se
cruzará y acabará por dirigirse a los
núcleos de los pares craneales del lado
contrario en donde se originó la vía. Al
igual que la vía piramidal, la geniculada
comienza en el Área 4.
En lo que se refiere a la neurona del
tronco encefálico o neurona motora inferior, cabe decir que posee características
similares a la segunda neurona de la vía piramidal, pues termina en el músculo efector y
en ella sinapta la primera neurona de la vía.
En relación con la vía geniculada, hemos de decir que existe un par craneal con
características especiales: el Facial o VII par. En el núcleo del facial, existen dos tipos
de neuronas funcionales: unas que conforman el Núcleo Facial Superior (1/3 superior)
y otras que configuran el Núcleo Facial Inferior (2/3 inferiores).El facial inerva
básicamente los músculos de la mímica además de los
músculos de los orificios naturales de la boca, sin
embargo, existe una distinción funcional, dado que los
músculos faciales por encima de los ojos presentan
inervación del núcleo facial superior. Además, la
característica más relevante, es que al núcleo facial
superior llegan neuronas y fibras de ambos hemisferios
124
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
corticales. Por ello, si existe una lesión podemos diagnosticar si es de primera o segunda
neurona: en una parálisis facial, si el paciente es capaz de arrugar los dos lados de la
frente, la lesión es de primera neurona, pues se afectaría la inervación de un lado, pero
seguiría funcionando con el otro. Por el contrario, si el paciente es incapaz de arrugar un
lado de la frente, concluimos que se trata de una afección en la neurona inferior.
APLICACIÓN CLÍNICA DEL CONOCIMIENTO DE LAS VÍAS MOTORAS
Existe una patología particular, que se denomina Hemiplejía Alterna que
consiste, por ejemplo en la parálisis motora de un lado del cuerpo y la parálisis facial
del lado contrario. La causa puede ser, por ejemplo, una lesión a nivel de la
protuberancia, que corta las vías descendentes piramidales antes de que se crucen
(provocan la parálisis del lado contrario del cuerpo) y afecta al núcleo del trigémino, en
una zona posterior al cruce de la vía geniculada (por lo que se paralizan los músculos
del lado contrario).
VÍAS EXTRAPIRAMIDALES
Aunque hasta el momento sólo hayamos visto vías descendentes que comienzan
en la corteza, hay que decir que existen vías descendentes motoras en la médula que no
proceden de la corteza, sino de estratos más bajos. Las más importantes son:
• Vía Tecto-espinal: existen, en realidad, dos vías tectoespinales, que se
sitúan, una al lado de la vía piramidal directa (vía tecto-espinal anterior) y
otra al lado de la vía rubroespinal (vía tectoespinal lateral). La vista y el
oído generan una serie de reflejos, y esta vía se encarga de recoger
información de los tubérculos cuadrigéminos y regular estos reflejos.
• Vía Rubro-espinal (núcleo rojo): tiene la función de inhibir los reflejos
extensores, sobre todo de las extremidades superiores (es decir, inhibe
movimientos antigravitatorios). Se sitúa, en la médula, por encima de la vía
cruzada.
• Vías del Retículo-espinal: existen también dos fascículos de la vía retículo-
espinal, uno anterior y uno lateral. El retículo espinal anterior suele venir de
la formación reticular del puente, y el retículo espinal lateral proviene,
normalmente, del bulbo. Van a actuar sobre el asta anterior, uno inhibiendo
125
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
los movimientos de extensión (el fascículo lateral), y el otro facilitándolos
(el fascículo anterior).
• Vías Vestíbulo-espinales (compuesta por fibras que los núcleos vestibulares
envían a la médula): también encontramos dos fascículos, uno anterior y otro
lateral, cuya función será facilitar los reflejos de equilibrio relacionados con
la gravedad y nos permite, así, mantenernos de pie sin gran esfuerzo.
• Fibras olivo-espinales: esta vía se situaría por delante del resto de vías, pero
para muchos autores, no existe en el ser humano (fue descubierta en gatos)
Todas estas fibras van a dar ramas para las interneuronas del asta anterior, de
forma que conseguirán regular el sistema motor en cada momento, en función de las
necesidades exigidas.
El conocimiento de la existencia de estas vías permite comprender las dos
posturas típicas de un paciente comatoso:
1. Postura de rigidez de decorticación: lesión por encima del núcleo rojo, por
lo que la persona está en flexión (los reflejos de extensión son inhibidos).
2. Postura de rigidez de cerebración: lesión por debajo del mesencéfalo. Se
afecta el núcleo rojo, por lo que los reflejos extensión no son inhibidos, y por
lo tanto, el paciente aparece en posición de extensión.
126
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VÍAS ASCENDENTES DE LA MÉDULA ESPINAL (SENSITIVAS)
Las vías ascendentes de la médula espinal son, principalmente, sensitivas y se
sitúan en sitúan en el cordón posterior y en el cordón antero-lateral. Esta distinta
distribución permite que se afecten unas mientras las otras permanecen intactas, dando
lugar a unas patologías específicas que cursan con una Disociación de la Sensibilidad.
Las vías principales que van a dirigirse al encéfalo a través del cordón posterior
de la médula son las que a continuación presentamos:
a) Vía tactil-epicrítica: es la sensibilidad del tacto de la piel que permite
distinguir dos puntos cercanos. Se mide, en clínica, con un compás (se van
juntando las puntas del mismo, hasta que el paciente siente sólo una punta).
Este tipo de sensibilidad no es igual en todas las regiones del cuerpo
humano.
b) Vías propioceptivas conscientes: sensibilidad propia del aparato locomotor,
que podemos sentir sin percibirla con los órganos de los sentidos. Cuando
examinamos la vía propioceptiva, debe hacerse privando al paciente de la
vista.Existen dos tipos de vías propioceptivas conscientes:
- Estática: identifica la situación de músculos y articulaciones.
- Dinámica: reconoce el movimiento de estructuras locomotoras.
c) Vías de la sensibilidad vibratoria: se trata de una variante sensitiva que se
explora mediante el empleo de un diapasón, que se sitúa sobre un elemento
óseo. Si el paciente es capaz de percibir la vibración, posee sensibilidad
vibratoria.
En cuanto a las vías que discurren por el cordón ántero-lateral de la médula,
podemos decir que son las siguientes:
a) Vías de la sensibilidad térmica: permiten el reconocimiento de cambios de
temperatura. Se exploran con pequeños tubos rellenos de agua a distinta
temperatura. Las vías de la sensibilidad térmica constituyen, junto con las
vías del dolor, lo que se ha denominado como Sensibilidad Termo-Algésica.
b) Vías de la sensibilidad al dolor: se encargan de identificar estímulos dañinos
para poder elaborar una respuesta adecuada frente a ellos.
c) Vías tactil-protopáticas: es la vía encargada del tacto “burdo”. Se testa con
un algodón sobre la piel. Si el paciente posee sensibilidad tactil epicrítica,
127
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
poseerá también la tactil-protopática, ya que ésta está incluida en la primera.
Por ello, la prueba de la sensibilidad tactil-protopática no es muy frecuente,
pues con frecuencia se realiza sólo la prueba para comprobar la sensibilidad
tactil epicrítica. También se denomina vía espino-talámica anterior.
d) Vías de la sensibilidad propioceptiva incosciente: actualmente, no existe
ningún método clínico que permita la exploración de estas vías.
VÍAS DE LA SENSIBILIDAD DEL CORDÓN POSTERIOR
Cualquier vía de la sensibilidad estará formada, en primer lugar, por los
receptores de los estímulos. Los receptores son transductores que cambian una energía
de cualquier tipo (eléctrica, mecánica, química,….) en impulsos nerviosos. Estos
receptores envían los impulsos nerviosos formados a través de una cadena de neuronas
hacia unos centros primarios de la sensibilidad que, como norma general, se encuentra
en la corteza (cuando nos referimos a la sensibilidad consciente). Estos centros
primarios enviarán la información a unos centros secundarios o de asociación, cuya
función es la interpretación del estímulo.
La diferencia entre las vías tactil-epicrítica, propioceptiva consciente y
sensibilidad vibratoria reside en los receptores de las tres vías. De esta manera, los
receptores de la sensibilidad tactil-epicrítica van a ser de muchos tipos, algunos de ellos
encapsulados, compuestos por fibras nerviosas que poseen una cápsula a su alrededor
(como los Corpúsculos de Meissner o de Ruffini). Receptores táctiles sin cápsula son,
por ejemplo, los que componen el complejo de Merckel (células sensitivas que entran en
contacto directo con una terminación nerviosa en la que descarga un impulso eléctrico
produciendo un estímulo). En cuanto a los receptores de la sensibilidad vibratoria, cabe
decir que van a emplear los mismos que la tactil-epicrítica, pero sólo los encapsulados
situados a mayor profundidad en la piel (cuanto más profundos estén los receptores,
más capas de cápsula poseerán).
La primera neurona de una vía sensitiva es casi siempre una Neurona
Ganglionar, situada en un ganglio sensitivo y que va a enviar la información,
directamente a través de los cordones posteriores, a una segunda neurona situada en el
bulbo (Neurona Bulbar). Esta segunda neurona conecta con una tercera que se encuentra
en el tálamo óptico y que se denomina Nuerona Talámica. Esta última va a enviar la
información a la corteza primaria, que a su vez la dirigirá hacia la corteza de asociación.
128
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La primera neurona ganglionar penetra con su axón por la zona marginal de
Lissauer sin sinaptar en ella, y asciende directamente por el cordón posterior hacia el
bulbo. No obstante, envía unas ramas colaterales a la médula, cuya función es la
filtración del dolor (pero nada tiene que ver
con la vía de la sensibilidad típica del
cordón posterior). Cuando el axón de la
neurona llega al bulbo, sinaptará con
neuronas del núcleo de Goll o de Burdach.
El núcleo de Goll se encarga de la
sensibilidad del cuerpo por encima del
dermatoma T6, mientras que el de Burdach
va a encargarse de la sensibilidad por debajo
de dicho dermatoma. Las neuronas que
parten de los susodichos núcleos de Goll y
Burdach se cruzan a nivel del bublo y
forman el Lemnisco Interno de la Cinta de
Reil o Cinta de Reil Medial. Posteriormente,
estas neuronas se dirigen al tálamo óptico, a
un lugar específico que se conoce con el nombre de Núcleo Ventro-Lateral Posterior.
Ya desde ahí, las neuronas proyectan sobre la corteza cerebral, en forma de radiaciones
talámicas. Concretamente, llegan a la circunvolución parietal ascendente. Si
representamos gráficamente las distintas áreas de esta circunvolución con las estructuras
corporales a las que inerva, obtendremos un “Homúnculo Sensitivo”, similar al
Homúnculo de Peinfield.
CORTE DE MESENCÉFALO
CORTE DE BULBO RAQUÍDEO
129
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
A nivel del bulbo, la vía sensitiva aparecerá por detrás del sistema motor, y
existirá una decusación sensitiva, fruto del cruce de las vías procedentes de los núcleos
de Goll y Burdach. En el puente o protuberancia, la vía sensitiva se va a localizar
también por detrás del sistema motor. A nivel mesencefálico, la Cinta de Reil Medial va
a ir por detrás de la sustancia negra.
En lo que se refiere a la sensibilidad propioceptica consciente (y también la
incosciente), cabe decir que va a seguir la misma vía que hemos explicado pero, como
ya hemos dicho, los receptores van a ser diferentes. La sensibilidad propioceptiva
empleará, pues, todos los receptores
táctiles (por lo que éstos se incluyen
indirectamente en la propiocepción).
Además de dichos receptores, todas las
cápsulas de las articulaciones poseen
unas terminaciones que intervienen en la
propiocepción. Finalmente, existen unos
receptores específicos de la propiocepción que son los Husos Neuromusculares de
Khüne y los Órganos Neurotendinosos de Golgi, que ahora explicaremos con más
detalle debido a su importancia clínica.
HUSOS NEUROMUSCULARES
Los husos neuromusculares son órganos fusiformes compuestos por fibras
musculares (más concretamente por fibras intrafusales). Estos corpúsculos fusiformes
están formados por una cápsula fibrosa del huso sujeta, por sus extremos, a las fibras
musculares normales de trabajo. De esta manera, estas estructuras se encuentran al lado
de un músculo estriado normal, y en su interior contienen Fibras musculares de la Bolsa
o Saco Nuclear (adaptadas a cambios rápidos), donde los núcleos se sitúan en la zona
central, formando un acúmulo nuclear. Igualmente, pueden contener otras fibras en las
que los núcleos se encuentran formando una cadena central, por lo que reciben el
nombre de Fibras musculares de la Cadena Nuclear (adaptadas a cambios lentos). En
ambos casos, las estriaciones se encuentran en los extremos, de forma que las partes
contráctiles de estas fibras se sitúan en las prolongaciones.
El hecho de que las fibras intrafusales se contraigan únicamente en los extremos
es tremendamente importante para la función del huso, pues presenta unas
130
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Terminaciones Sensitivas Ánulo-espinales, que van a acabar en las astas posteriores de
la médula espinal. Estas terminaciones hacen que se disparen las señales cuando se
estira el núcleo como resultado de la contracción de los extremos de las fibras
intrafusales.
Como observamos, el huso neuromuscular es, pues, un receptor del
estiramiento muscular, que actúa como un sistema de protección del músculo,
evitando extensiones excesivas del mismo. La inervación motora de esta zona la van a
realizar las motoneuronas gamma, que provienen del asta anterior de la médula, y se
dirigen únicamente a los extremos de las fibras contráctiles. Así, cuando los extremos se
contraen, las bolsas y cadenas nucleares se estiran, disparando las señales sensitivas. De
esta manera, se va a informar al asta posterior del estiramiento el músculo. No obstante,
la función de los husos no termina aquí, pues las neuronas que componen las
terminaciones ánulo-espinales envían colaterales que llegan hasta el asta anterior de la
médula y que harán sinapsis con una motoneurona alfa, activándola y provocando la
contracción del músculo estriado adyacente al huso. De la misma forma, mediante una
interneurona, se actúa sobre los antagonistas del músculo que se tiene que contraer para
que se relajen y pueda tener lugar el movimiento.
131
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El sistema de los husos neuromusculares sienta las bases del reflejo ósteo-
muscular u ósteo-tendinoso. Así, cuando se golpea un tendón, este se extiende
rápidamente, por lo que el músculo al que pertenece se va a contraer rápidamente
gracias a la acción de los husos neuromusculares.
El sistema gamma también posee otra función, pues hace que la contracción de
un músculo sea continua, sin elementos bruscos. Esto se debe a que, antes de iniciar la
contracción muscular, el sistema gamma ya se pone en acción, y va contrayendo los
extremos de las fibras intrafusales para modular la contracción muscular.
Otro aspecto relacionado con el sistema de los husos musculares, es el hecho de
que permite la contracción de la musculatura de trabajo mediante la activación directa
de la motoneurona gamma. Este es el sistema empleado, normalmente, para crear lo que
se conoce como Tono Muscular, es decir, para mantener grado mínimo de contracción
muscular que permite mantener una postura de forma automática.
Una vez conocido el funcionamiento de los husos neuromusculares, podemos
comprender la Espasticidad, una tensión constante del músculo que aumenta con la
extensión, resultado de una sobreestimulación del circuito gamma. Por ello en las
parálisis espásticas, en las que se afecta la primera neurona, se mantiene la espasticidad
y el tono muscular, pues sobre la motoneurona gamma van a actuar una serie de
fascículos extrapiramidales. Además, cabe especificar que, en las lesiones de primera
neurona, existe un aumento de la acción del circuito gamma, por lo que se producirá
hiperreflexia. Esto se debe a que las fibras que acompañan a la primera neurona son
inhibidoras del circuito gamma (disminuyen el reflejo).
Como hemos visto hasta el momento, los husos neuromusculares son los
responsables de conocer el grado de estiramiento de un músculo, pero es necesario
también, para la propiocepción, conocer la fuerza muscular, lo que se consigue con los
órganos neurotendinosos de Golgi.
ÓRGANOS NEUROTENDINOSOS DE GOLGI
Se trata de unos receptores de tracción. Cuando el músculo se contrae,
transmiten la fuerza a los tendones. En dichos tendones (sobre todo en las uniones
miotendinosas), aparecen los órganos neurontendinosos de Golgi, que se activarán y
dispararán estímulos como respuesta a la contracción y a la fuerza que el músculo
transmite al tendón. Así, cuando se contrae el músculo, el órgano envía información al
132
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
sistema propioceptivo y a través de una interneurona inhibidora dará órdenes a la
neurona alfa del asta anterior para que el músculo se relaje. Como se puede deducir, se
trata de un sistema defensivo del músculo frente a la contracción, hasta el punto de
que, cuando una persona no tiene el sistema propioceptor en condiciones, aumentarán
considerablemente el número de lesiones musculares.
Los sistemas propioceptivos son también los responsables de movimientos
contrariados (contraer un músculo y relajar su antagonista). De hecho, existen algunos
reflejos que tienen su base en estos sistemas, como es el caso de los que a continuación
presentamos:
• Reflejo flexor polisináptico o de triple retirada: consiste en la flexión de la
cadera, rodilla y tobillo. Se trata de una vía complicada en la que se recoge
información de la vía del dolor, y se envía a varios segmentos medulares, que
provocarán la contracción y la retirada del estímulo doloroso. Este reflejo se
vuelve patológico cuando se activa ante cualquier estímulo cutáneo. Esto ocurre,
por ejemplo, en lesiones de primera neurona, dado que las fibras que inhiben
este reflejo no funcionan.
• Reflejo extensor cruzado: consiste en la flexión de una extremidad como
consecuencia de una extiensión excesiva de la otra. En condiciones normales,
este reflejo está muy inhibido el en hombre. No obstante, en algunos animales
está muy desarrollado, hasta el punto de que ciertas especies pueden llegar a
andar de forma refleja.
133
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VÍAS DE LA SENSIBILIDAD DEL CORDÓN ANTERO-LATERAL
Se trata de una vía situada en la región anterior y lateral de la médula, cuya
función principal es la percepción de la temperatura y el dolor (Vía Termoalgésica).
Además de ésta, por el cordón lateral van también las vías tactil-protopática y de la
sensibilidad propioceptiva inconsciente.
CONCEPCIÓN MÉDICA DE DOLOR
Submodalidad de sensación somática de signo de alarma, con función
protectora, que nos avisa de que una lesión debe ser evitada. Puede ser definido también
como cualidad urgente y primitiva con aspectos afectivos y emocionales en su
percepción, dado que, cuando decimos que el dolor es una sensación, decimos que se
trata de una percepción personal. El dolor es, igualmente, una percepción desagradable
asociada con lesión tisular presente o posible, producto de la abstracción y elaboración
por parte del cerebro de una señal sensitiva.
El dolor tiene muchos matices en función de la persona que los siente, de forma
que, como médicos, nos encontraremos frecuentemente los siguientes tipos de dolor:
• Pinchazos
• Quemazón.
• Dolorimiento.
• Picaduras.
• Dolor sordo.
• Como corriente eléctrica.
TIPOS DE DOLOR
Para poder comprender el dolor, se ha clasificado en varios tipos en función de
diferentes aspectos. Algunas de las clasificaciones más frecuentes son las que
presentamos a continuación.
Según el origen del estímulo doloroso, podremos diferenciar tres tipos
fundamentales de dolor, que son:
134
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Dolor Somático: en este caso, existe un receptor que transforma impulsos
mecánicos, físicos y eléctricos en impulsos nerviosos, frente a los que se
elabora una respuesta dolorosa.
• Dolor Neurológico: el origen de dolor se sitúan en alguna de las vías del
sistema nervioso. No procede, por tanto, de un receptor, y se va a afectar la
zona correspondiente que sea inervada por la raíz dañada.
• Dolor Psíquico: aparece en algunas patologías psíquicas, y para él no existe
un origen objetivo.
En relación con la significación clínica del dolor, hemos de decir que éste va a
poder claisificarse en:
• Dolor Persistente: es característico de muchas enfermedades. Se trata de un
dolor inútil, pues pierde su función de alarma una vez que el estímulo dañino
cesa. A su vez, el dolor persistente puede ser de dos tipos:
- Nociceptivo
- Neuropático
• Dolor Crónico: no presenta ninguna función útil y sólo sirve para amargar la
vida al paciente.
Además de los que hemos señalado, existen otros tipos de dolor que siempre
implican problemas médicos y que son originados en lugares diferentes al lugar en que
el paciente percibe el dolor. Estos tipos de dolor son:
• Dolor Irradiado: se trata del dolor resultante de la irritación de un nervio (es
un dolor neuropático). Implica dolor en todo el territorio inervado por el
nervio o raíz nerviosa correspondiente.
• Dolor Diferido: se trata de un dolor más complicado, dado que suele ser
visceral. Se trata de un dolor sordo y difuso que se va a sentir diferentes
partes del cuerpo en función de la víscera afectada:
- El corazón: puede sentirse en cualquier parte del hemitórax izquierdo
y en la extremidad superior izquierda.
- El estómago: se sentirá en el espación subxifoideo.
- La vesícula: el dolor será percibido por debajo de las costillas.
- Los riñones: dolor que pasa a las ingles a nivel de la cresta ilíaca, y
que se siente también por un costado.
- El diafragma: se siente en el hombro izquierdo.
135
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
DOLOR SOMÁTICO
Se trata de un dolor que comienza en unos receptores nociceptivos que son, en
su mayoría, terminaciones nerviosas libres. Están en todos los órganos y todas las partes
del cuerpo salvo en el sistema nervioso central, que no posee, por tanto, dolor somático
(sólo lo habrá neuropático). La neurona ganglionar de la vía llega hasta el asta posterior
de la médula, y desde ahí, por varias vías, llega a los centros procesadores.
RECEPTORES DEL DOLOR
Suelen ser terminaciones nerviosas libres. Existen varios tipos de receptores
algésicos: térmicos, mecánicos, químicos, polimodales y receptores silentes (capaces de
recibir estímulos que producen dolor, pero que no están dedicados únicamente a esta
vía). Cabe decir que el principal receptor en las terminaciones libres es la capsaicina.
Igualmente, es de importancia señalar el hecho de que las fibras A-delta (Aδ)
son las transmisoras del dolor agudo, mientras que las fibras C son las responsables del
dolor lento.
En cuanto a la recepción del dolor, existen una serie de estados patológicos que
reciben nombres específicos en función de la sintomatología. Los más destacados son
los que presentamos a continuación:
• Alodinia: se produce dolor con estímulos inocuos.
• Hiperalgesia: respuesta excesiva a estímulos nocivos. Esta patología se
produce frecuentemente en el organismo, ya que después de un traumatismo
fuerte, en la zona afectada, permanecen una serie de neurotransmisores y
neuromoduladores que mantienen la zona en un estado de “dolorimiento”
durante un período de tiempo más o menos largo (bajando el umbral de los
receptores).
• Anestesia dolorosa: dolor en ausencia de sensación (el paciente no siente
nada que no sea dolor).
NEURONAS MEDULARES RELACIONADAS CON LAS VÍAS DEL DOLOR
Las neuronas medulares que pertenecen a las vías del dolor van a poseer una
serie de características específicas, tal y como a continuación procedemos a enumerar:
136
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Están situadas en el asta posterior de la médula en las láminas I, II, III, IV y
V de Rexed. No obstante, en la sustancia gelatinosa de Rolando se realizará
únicamente un filtro de las señales dolorosas.
• El principal neurotransmisor de dichas neuronas es el glutamato.
• Los neuropéptidos, incluida la sustancia P, potencian y prolongan la acción
del glutamato. Un estímulo importante del dolor son los ácidos grasos de
cadena ligera (liberación de los ácidos grasos de las membranas por necrosis
tisular), que actúan a nivel del receptor y a nivel medular.
• La liberación de ciertas sustancias químicas produce sensibilización a nivel
central y periférico. Algunas de dichas sustancias son la bradicinina, las
prostaglandinas, los leucotrienos, la acetilcolina, la serotonina, y la
sustancia P (inflamación neurógena).
• La inducción de la transcripción de genes de tipo c-FOS en las neuronas del
asta posterior produce neurotransmisores sensibilizadores.
TRANSMISIÓN DE LAS SEÑALES NOCICEPTIVAS
Existen, en el cuerpo humano varios haces de fibras ascendentes cuya función
principal es la transmisión de la información nociceptiva. Algunas de ellas son:
a) Haz espinotalámico: se encarga de transmitir el dolor agudo. La segunda
neurona se cruza con la médula espinal y asciende por el cordón lateral hasta el
tálamo óptico. Del tálamo, se dirige directamente a la corteza. Esta vía se conoce
también con el nombre de neo-espinotalámica.
b) Haz espinorreticular: para el dolor sordo, y hace sinapsis en la formación
reticular antes de dirigirse al tálamo. Es una vía un poco más lenta y se conoce
como paleo-espinotalámica.
c) Polisináptico difuso: se sabe que es una vía cruzada, pero que está formada por
múltiples neuronas situadas en el tronco del encéfalo, conectando con la
formación reticular y llegando hasta el tálamo óptico. Se encarga del dolor
visceral.
d) Haz espinomesendefálico.
e) Haz cervicotalámico.
f) Haz espinohipotalámico.
137
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CENTROS PROCESADORES Y RECEPTORES
Al igual que existen diversos haces de fibras que transmiten la información
nociceptiva, van a existir una serie de núcleos, ya en el sistema nervioso centrales, que
se van a encargar de procesar la información que llega desde estos haces:
• Núcleos del tálamo óptico:
a) Núcleo Ventrolateral Posterior: a él llega la vía neo-espinotalámica.
b) Núcleos reticulares talámicos: al que pertenecen el resto de las vías.
c) Grupo nuclear interno.
CORTEZA CEREBRAL
En lo que se refiere a la corteza cerebral, las vías nociceptivas van a estar en
relación con una serie de áreas concretas, que son las siguientes:
a) Corteza Somatosensitiva: contribuye a la localización del estímulo doloroso.
Se sitúa en la circunvolución parietal ascendente, y de aquí parten fibras para
el gyrus cínguli (perteneciente al sistema límbico).
b) Circunvolución singular: conexión con el sistema límbico. Aquí el dolor se
hace agradable o desagradable, en relación con la persona que lo recibe.
c) Ínsula: integra los componentes sensitivos, afectivos y cognitivos del dolor.
CONTROL CENTRAL DEL DOLOR
Para explicar el control central debemos hacer referencia directa a la Teoría de la
Puerta de Entrada del Dolor (Gait Control). Esta teoría se basa en los sucesos que
ocurren tras una estimulación eléctrica transcutánea (estimulación de cordones
posteriores), dado que, como hemos visto, la vía tactil-epicrítica envía colaterales al
asta posterior, que sirven para controlar la entrada del dolor. Esta teoría, de Melzack y
Wall explica muchos aspectos del control del dolor, y sostiene que, si se estimulan las
fibras gruesas (Fibras A), se cierra la puerta del dolor, mientras que si son las fibras C
las estimuladas, además de transmitirse la señal nociceptiva, se va abrir la puerta del
dolor. La función analgésica de los masajes, por ejemplo, radica en que son estímulos
sensitivos que cierran la puerta del dolor, estimulando las fibras A (aunque también se
producen endorfinas endógenas que actúan como analgésico). En esta teoría se basa
138
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
también el funcionamiento de los TENS, sistemas de estimulación transcutánea del
sistema nervioso, que se encargan de estimular las fibras A para cerrar la puerta del
dolor.
Las vías córtico-espinales y retículo-espinales mandan colaterales a la médula
para inhibir las vías del dolor. Se trata de sistemas inhibidores descendentes que se
activan con una estimulación eléctrica del III ventrículo y el acueducto de Silvio.
Algunos péptidos opioides endógenos, como son las encefalinas, B-endorfinas y
dinorfinas, producen una sensación de bienestar y eliminan el dolor. Sustancias de este
tipo se producen, por ejemplo, con el ejercicio físico. El estrés puede producir también
analgesia por mecanismos opioides y no opioides.
TIPOS DE ANESTESIA
Como conclusión, hemos de decir que existe una forma clínica de producir
analgesia temporal gracias a la inyección de ciertas sustancias en el paciente: es la
Anestesia. Los tipos más importantes de anestesia son los que presentamos a
continuación:
• Anestesia local: inyectamos el anestésico en una zona concreta.
• Anestesia Intravascular: el anestésico se introduce en un vaso sanguíneo.
• Simpatectomía visceral
• Anestesia Epidural: inyectamos el anestésico en el espacio epidural.
• Intratecal: el anestésico se introduce dentro del saco de la duramadre.
• Infiltración ganglionar: se anestesia un ganglio.
APLICACIONES CLÍNICAS
Para comprobar el estado de la vías de la sensibilidad, hemos de explorar los
distintos dermatomas corporales, de forma que podamos determinar si existe afección
en alguna de las vías sensitivas que a ellos llegan.
Cuando un paciente presenta las vías del cordón posterior normales y la vía
termoalgésica afectada, decimos que existe una Disociación Siringomiélica. En el caso
de la Disociación Tabética, por el contrario, existe lesión en alguna de las vías de los
cordones posteriores.
139
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Para comenzar con el estudio de la morfología interna del tronco encefálico,
señalaremos una serie de homologías entre éste y la médula espinal que nos permitan
comprender, así, las distintas partes que lo componen. Las homologías más importantes
a las que hacemos referencia son:
• Tanto de la médula espinal como del TE salen una serie de nervios en
parejas, que serán los pares raquídeos y los pares craneales.
• Ambas estructuras van a estar constituidas por sustancia gris y sustancia
blanca.
No obstante, existen una serie de características en las que médula y TE se
diferencian, y serán éstas las que definan cada una de las partes. Las más importantes
son:
• Presentan distinta disposición de la sustancia gris. En la médula, la sustancia
gris se presenta formando un bloque único, en la zona media, rodeada de la
sustancia blanca. En el TE, por el contrario, la sustancia gris se distribuirá de
forma difusa agrupándose en acúmulos denominados Núcleos y va a estar
“sumergida” en una gran cantidad de fibras.
• Los elementos de asociación entre zonas de materia gris en la médula espinal
son muy poco frecuentes, mientras que en el TE, la relación directa entre los
núcleos y los nervios es muy escasa (todo lo contrario a la médula), ya que la
mayoría de núcleos y centros neuronales se van a relacionar y coordinar con
otros centros (y no con fibras nerviosas)
DISTRIBUCIÓN Y ELEMENTOS PRINCIPALES DE LA CONFIGURACIÓN
INTERNA DEL TE
La división que a continuación presentamos se trata de una clasificación clásica
con una finalidad únicamente didáctica, pues hoy día se saben que algunos de esos
núcleos se superponen, y por tanto no pueden ser definidos independientemente.
Sabiendo esto, contemplamos la siguiente clasificación a la hora de estudiar la
morfología interna del tronco encefálico:
140
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Núcleos Segmentarios: son los núcleos en los que se sitúa el origen de los pares
craneales y reciben el nombre de segmentarios porque proceden de distintos
segmentos embriológicos.
• Núcleos suprasegmentarios: se trata de núcleos superiores que van a coordinar a
los núcleos segmentarios. Son los tubérculos cuadrigéminos, el núcleo rojo y la
sustancia negra.
• Núcleos intersegmentarios: son núcleos que se agrupan formando centros de
asociación. Dentro de este grupo de núcleos podemos establecer una jerarquía,
dado que no todos son iguales (a medida que ascendemos en el SNC aumenta el
nivel en la jerarquía). La sustancia reticular es un ejemplo claro de este tipo de
núcleos.
DISTRIBUCIÓN Y ELEMENTOS PRINCIPALES DE LA CONFIGURACIÓN
INTERNA DEL TE
NÚCLEOS SEGMENTARIOS
El origen embriológico de los pares craneales tiene lugar en los somitas cefálicos
por un lado, y en los arcos braquiales por el otro. Este origen puede deducirse del hecho
de que, a partir de los somitos cefálicos se origina la musculatura extraocular y la
musculatura lingual, mientras que el resto de estructuras inervadas por los pares
craneales tienen su origen en los arcos branquiales.
Los pares craneales que derivan de los somitos embrionarios pueden clasificarse
en dos grandes grupos:
• Preópticos: inervan los músculos extraoculares y presentan, por tanto, una
función que es exclusivamente motora. Son los pares craneales III, IV y VI.
• Occipitales: Inervan los músculos linguales (XII par craneal).
Los arcos braquiales, por lo tanto, van a dar origen a la musculatura del cuello y
la cabeza, y cada arco braquial tendrá relación con un par craneal y con la musculatura
que de él deriva.
• Primer arco: mandibular (V par)
• Segundo arco: hioideo (VII par)
• Tercer arco: da lugar al IX par.
141
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Cuarto arco: hiotiroideo
Son homólogos a la sustancia gris de la médula espinal, y presentan una
distribución uniforme, formando columnas incompletas en el TE. Existen, por ello, tres
columnas que tienen asignadas funciones nerviosas diferentes:
1. Columna Motora: situada en la línea media. Presenta el origen de los pares
somíticos XII, VI, IV y III y de los pares braquiales X, XI, IX VII y V.
2. Columna Vegetativa: pegada a la cavidad del cuarto ventrículo, rodeándola.
Existen aquí origen para los pares X, IX, VII y III.
3. Columna Sensitiva: situada por fuera de la cavidad del cuarto ventrículo. Los
pares craneales que presentan origen en esta columna son el X, el IX, el VII, el
V y el VIII.
Como hemos observado, será el suelo del cuarto ventrículo el que va a
determinar la distribución que acaba de ser explicada. Cabe decir, no obstante, que en
todo el TE aparecen una serie de fibras cruzándose.
Como resultado, podemos decir que los núcleos segmentarios dan lugar a doce pares
craneales (I-XII) que se enumeran según emergencia del cráneo, y que se van a
clasificar, desde un punto de vista embriológico en:
a) Somíticos: se trata de pares craneales cuya función es exclusivamente motora.
Éstos, a su vez, se dividen en dos subtipos:
- Preópticos.
- Occipitales.
b) Braquiales: derivan de los arcos braquiales y no exclusivamente motores.
LOCALIZACIÓN DE LOS NÚCLEOS SEGMENTARIOS EN EL TE
LOCALIZACIÓN DE LOS NÚCLEOS
PARES CRANEALES Bulbo Protuberancia Mesencéfalo
III PAR (Motor Ocular Común)
IV PAR (Patético o Troclearis)
V PAR (Trigémino)
VI PAR (Motor Ocular Externo)
142
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
VII PAR (Facial)
VIII PAR (Estatoacústico)
IX PAR (Glosofaríngeo)
X PAR (Vago)
XI PAR(Espinal)
XII PAR (Hipogloso)
III PAR, MOTOR OCULAR COMÚN
El núcleo del III par se sitúa en la parte más alta del mesencéfalo, cerca del
Tubérculo Cuadrigémino Superior y próximo al acueducto de Silvio. Se trata de un
núcleo exclusivamente motor, cuyos axones van a salir por delante del pedúnculo
cerebral, y que presenta forma de “H”, con interneuronas de coordinación (Núcleo de
Perlia) que actúan sobre los músculos de la convergencia ocular. A nivel de las partes
laterales del núcleo en forma de H, nos encontraremos con regiones en las que se
localizan el origen para la invervación de los distintos músculos extraoculares (de arriba
abajo):
a) Músculo Elevador del Párpado.
b) Músculo Recto Superior
c) Músculo Recto Interno
d) Músculo Recto Inferior
e) Músculo Oblicuo Menor.
En la parte más alta e interna (entre los dos núcleos motores), está asociado el
núcleo de Edinger-Westphal, centro vegetativo parasimpático que dará la inervación
parasimpática del músculo ciliar.
Entre el pie y la calota del TE,
nos encontramos con la Sustancia
Nigra, compuesta por dos porciones: la
Pars Compacta (más posterior) y la
Pars Reticulata (más anterior). En la
parte media de la calota aparece otro
núcleo denominado, por su coloración,
143
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Núcleo Rojo. Es una referencia para nosotros, pues será atravesado por los axones del
tercer par. El Núcleo del nervio motor ocular común estará también en relación con la
porción medial de la Cintilla de Reil y el Fascículo Longitudinal Lateral.
IV PAR, PATÉTICO O TROCLEAR
El núcleo del troclearis se sitúa en la parte inferior del mesencéfalo y es
exclusivamente motor. Se localiza en la parte posterior, con unas fibras que salen por el
Frenillo de la Válvula de Viussens. Las fibras del IV par pertenecen a la motoneurona
del lado contralateral
VI PAR, MOTOR OCULAR EXTERNO
El núcleo del VI par craneal se sitúa en el puente, en el suelo al IV ventrículo,
pegado a él a nivel de la Eminencia Teres. La Eminencia Teres es, de hecho, una
prominencia en el cuarto ventrículo que se origina cuando los axones del VII par rodean
al núcleo del VI par. Este núcleo enviará los axones hacia abajo y hacia fuera, de forma
que el origen aparente de las fibras del nervio motor ocular externo será el surco bulbo-
protuberancial. Este nervio se va a encargar de la inervación del Recto Externo
(músculo extrínseco del ojo).
144
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
XII PAR CRANEAL O HIPOGLOSO MAYOR
Va a presentar su origen en el suelo del cuarto ventrículo a nivel bulbar, más
concretamente en el ala blanca interna, y sus fibras saldrán a través del surco preolivar
(o surco del hipogloso). Se trata de un par craneal puramente motor.
V PAR O TRIGÉMINO
Es derivado del primer arco braquial del embrión. Se trata, por tanto, de un
nervio que ya no presenta únicamente funciones motoras, sino que también realizará
otras sensitivas.
En relación con las funciones motoras, cabe decir que el trigémino se relaciona
con los músculos masticadores, así como con la inervación del músculo martillo.
En lo que se refiere a las funciones sensitivas, hemos de comentar el hecho de
que se va a relacionar, principalmente, con la sensibilidad termo-algésica de la cara (ojo,
fosas nasales, fosas paranasales y duramadre).
Si realizamos un corte en la protuberancia, por encima del núcleo del facial, nos
vamos a encontrar con el núcleo motor del quinto par (a nivel de la calota de la
protuberancia). Igualmente, podremos
observar una parte de los núcleos
sensitivos del trigémino. No obstante,
hemos de aclarar que la sensibilidad
sensitiva del trigémino, en realidad, está
repartida por todo el tronco encefálico (en
el bulbo, la protuberancia e incluso llega hasta el mesencéfalo y los segmentos
medulares superiores).
De esta manera, existe un núcleo principal sensitivo a nivel de la protuberancia,
que va a enviar fibras ascendentes al mesencéfalo por un lado, y descendentes al bulbo
por el otro. Así, se configuran tres núcleos sensitivos diferentes para el trigémino:
• Núcleo de raíz transversa: también llamado núcleo principal, se localiza en la
protuberancia o puente. Este núcleo se especializará en la sensibilidad táctil
(principalmente discriminativa)
• Núcleo de raíz ascendente: se establece a nivel del mesencéfalo y se va a
especializar en la sensibilidad propioceptiva.
145
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Núcleo de raíz descendente: situado en la región del bulbo y especializado en
la sensibilidad termo-algésica. En lo que se refiere a este núcleo, las fibras
pueden llegar hasta los segmentos superiores de la médula, pero en cualquier
caso, enviará la información hacia el núcleo principal.
Esta distribución de la función sensitiva del trigémino a lo largo de todo el
tronco encefálico va a conllevar una serie de aspectos clínicos que son de especial
interés destacar:
• Dado que las fibras sensitivas se entremezclan a menudo en el TE, cuando
existe un dolor cervical, puede tener un origen cráneo-facial. Así mismo, un
dolor cráneo-facial puede presentar un origen cervical.
• El estudio del reflejo del cierre del párpado tras una leve estimulación
corneal es muy empleado para comprobar el nivel de conciencia de los
pacientes en coma.
VII PAR O NERVIO FACIAL
El origen de este par procede del segundo arco braquial (tirohioideo). Se trata de
un nervio mixto, con componente motor, sensitivo y vegetativo. Con respecto a su
inervación motora, va a estar relacionado con los músculos de la mímica, así como el
músculo del estribo, facilitando la acomodación auditiva. En cuanto a su función
vegetativa es de interés señalar que se encarga de la inervación de la glándula
lacrimonasal y de la salivar superior.
El núcleo motor se va a localizar en el tercio posterior del puente, mientras que
el sensitivo lo hará por delante y por dentro del núcleo del V par craneal, en la parte alta
146
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
del bulbo (a nivel del Núcleo Solitario del bulbo). Con respecto a la inervación
sensitiva, hemos de decir que puede ser general y especial. En cuanto a la inervación
especial, hemos de decir que estará relacionada con la recepción gustativa a nivel de la
“V lingual” (papilas caliciformes por delante de la V). La sensibilidad general se refiere
a la inervación sensitiva de regiones en general (desde una muela hasta una mejilla)
En relación con los núcleos vegetativos debemos señalar que se van a encargar
de la inervación de la glándula lacrimonasal (con el núcleo situado a nivel del suelo del
puente) y de la glándula salivar superior (Núcleo salivar superior).
IX PAR O GLOSOFARÍNGEO
Deriva del tercer arco braquial (tirohioideo). Es muy completo, pues tiene parte
motora, parte sensitiva y parte vegetativa (motora y sensitiva). En relación con su
función motora, va a inervar al músculo estilofaríngeo. Con respecto a su inervación
sensitiva, es preciso decir que posee una función sensitiva especial en relación con la
lengua para captar, principalmente, la sensación del sabor amargo. Esta función se sitúa
a nivel de la V lingual, en las papilas caliciformes. Igualmente, el glosofaríngeo recoge
la sensibilidad de la faringe y del istmo de las fauces.
En cuanto a sus funciones vegetativas, cabe comentar que pueden dividirse en
funciones vegetativas de estirpe motora y funciones vegetativas de estirpe sensitiva.
Así, por ejemplo, una función vegetativa motora estaría en relación con la glándula
parótida, mientras que una función vegetativa sensitiva estaría en relación con los
receptores del seno o cuerpo carotídeo (inervación sensitiva vegetativa interoceptiva).
La inervación vegetativa sensitiva interoceptiva consiste en que el cuerpo carotídeo, que
se encuentra a nivel de la bifurcación de la carótida primitiva, va a enviar información al
TE para comprobar el estado de la frecuencia cardíaca y la presión arterial, gracias a sus
quimiorreceptores y mediante el IX y X par. El nervio glosofaríngeo y el nervio vago se
comunican a través de un núcleo que se denomina Núcleo de Hering, al que llega la
información proveniente del seno carotídeo. Existen también otros receptores: los
corpúsculos carotídeos, que se sitúan a nivel de los capilares (entre la carótida interna y
la carótida externa), y que también van a estar conectados con el IX y el X par. Cuando
dicha información llega al TE, éste podrá modificar, en caso de que sea necesario, la
frecuencia cardíaca o la presión arterial.
147
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El núcleo del IX se sitúa en el bulbo, más concretamente en la parte superior del
mismo. Presenta distintos núcleos para sus distintas funciones:
a) Núcleo motor: lo constituye el Núcleo Ambiguo, situado en el centro de la
calota.
b) Núcleo sensitivo: es el Núcleo Solitario, que se localiza por detrás y por fuera en
relación con el núcleo ambiguo.
c) Núcleos vegetativos: Existen dos núcleos vegetativos, que son los que a
continuación exponemos:
- Núcleo salivar inferior (Núcleo redondo)
- Núcleo sensitivo dorsal.
X PAR, VAGO O PNEUMOGÁSTRICO
Procede del cuarto arco braquial, al igual que el nervio espinal (XI par). Tal y
como ocurre con el glosofaríngeo, se trata de un nervio motor, sensitivo y vegetativo
(sensitivo y motor).
En lo que se refiere a su función motora, hemos de decir que inerva la faringe.
En cuanto a la función sensitiva, va a dar inervación a la faringe y a una región que se
sitúa a nivel del Conducto auditivo externo y que se denomina “Zona de Ramsey-Hunt”.
Es ésta la parte más profunda del conducto auditivo externo y la concha. De este modo,
tenemos que su inervación motora la compartirá con el noveno par (núcleo ambiguo),
mientras que la sensitiva la compartirá con otros nervios (como es el caso de los pares
VII y IX).
148
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La inervación vegetativa motora corre a cargo de un núcleo de tremenda
importancia que recibe el nombre de Núcleo Cardio-reno-pneumo-entérico, situado por
debajo del Núcleo salivar inferior. Es moderador de la frecuencia cardíaca, desempeña
funciones a nivel renal, a nivel del aparato respiratorio, en relación con glándulas y
musculatura (traqueal, por ejemplo), y lleva a cabo diversas funciones en el aparato
digestivo. Esta inervación se sitúa a nivel de la faringe, esófago, estómago, hígado,
intestino delgado, y parte del intestino grueso, en relación con la musculatura lisa y la
membrana.
La inervación vegetativa sensitiva va a compartir inervación con el
glosofaríngeo. Dicha función se sitúa en los núcleos redondo y el sensitivo dorsal, que
son los encargados de recoger las sensaciones de la vía sensitiva vascular del seno
carotídeo y los corpúsculos carotídeos. Se situarían entre la parte superior del bulbo y el
puente.
XI PAR CRANEAL O ESPINAL
Es un par craneal que deriva de los arcos braquiales cuarto y quinto. Es
exclusivamente motor e inerva toda la musculatura de la laringe excepto el crico
aritenoideo posterior. Dará inervación, también, para los músculos
esternocleidomastoideo y trapecio. De esta manera, puede ya deducirse la existencia de
dos núcleos:
• Núcleo a nivel bulbar: que se localizaría en el tercio inferior del ambiguo.
• Núcleo a nivel medular: en la parte lateral del asta anterior, concretamente a
nivel del núcleo céfalo-giro. Por una parte, este núcleo céfalo-giro daría un
nervio externo para el esternocleidomastoideo y el trapecio y otro más interno
que inerva la musculatura laríngea.
VIII PAR CRANEAL O ESTEATOACÚSTICO
Es únicamente sensitivo, y tiene una parte coclear y otra vestibular. La parte
coclear tiene su origen real en el Tubérculo Acústico, situado en la parte inferior del
puente o protuberancia y va a estar regida por dos núcleos: el Núcleo Dorsal y el Núcleo
Coclear Ventral.
149
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La parte vestibular se encuentra en el ala blanca externa, y estará regida por los
siguientes núcleos:
• Núcleo de Betcherew.
• Núcleo de Schbalbe.
• Núcleo de Deiter.
• Núcleo de Roller.
• Núcleo de Lewandosky.
NÚCLEOS SUPRASEGMENTARIOS
Se trata de una serie de núcleos cuya función principal será la coordinación de
los núcleos segmentarios o de los pares craneales. Los núcleos suprasegmentarios más
importantes son los que a continuación presentamos y explicamos:
1. Tubérculos cuadrigéminos: existen cuatro tubérculos cuadrigéminos, divididos
dos a dos en superiores e inferiores, y que presentan funciones distintas:
- Tubéculos Cuadrigéminos Superiores: están distribuidos a través de
láminas de sustancia gris y sustancia blanca superpuestas. A ellos llegan
ramas colaterales de la vía óptica, de forma que su función principal será
ña coordinación o regulación de reflejos de la visión.
- Tubérculos Cuadrigéminos Inferiores: no presentan la misma estructura
laminar, y la función que desempeñarán será la emisión de ramas
colaterales en relación con la vía auditiva. Lo hacen mediante un haz que
se conoce como Tecto-espinal.
2. Núcleo Rojo: se sitúa delante de los tubérculos cuadrigéminos. En el núcleo
rojo, nos encontramos con dos partes: una porción magnocelular situada a nivel
inferior, y una porción parvocelular, situada a nivel superior. La función
principal de este núcleo es que interviene en la ejecución de movimientos
voluntarios a través del haz Rubro-espinal, dado que inhibe la vía piramidal.
3. Sustancia nigra: actúa regulando las vías. Presenta dos partes con funciones
diferentes, que son la pars compacta y la pars reticulata (aunque ambas modulan
la vía extrapiramidal):
- Pars Reticulata: se conectará (principalmente) con el pálidum a través
de aferencias y mediante sinapsis de un neurotransmisor específico, que
es el GABA (conexiones gabaérgicas).
150
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
- Pars compacta: establece relaciones con el estriado mediante un
neuromodulador específico, la Dopamina.
4. Oliva bulbar: se sitúa en la cara lateral del bulbo. Por dentro, la oliva presenta
unas paredes plegadas llenas de fibras, dado que a ella llegan gran cantidad de
fibras procedentes de otras localizaciones. La oliva está cubierta por una cápsula
que, si se abre, se observa que a ella llegan aferencias de haces espino-olivares y
que de ella parten fascículos olivo-espinales. Este núcleo se relaciona también
con la vía extrapiramidal.
5. Paraoliva: localizada en el puente, detrás de las fibras transversales. Presenta
relación con los reflejos auditivos.
NÚCLEOS INTERSEGMENTARIOS
Se trata de grupos nucleares que se asocian y que están regulados con una cierta
jerarquía, formando un sistema a lo largo del tronco-encéfalo que se conoce con el
nombre de Sistema Reticular. Éste puede definirse como una red tridimensional de
grupos de interneuronas o de grupos reticulares que, desde el tronco-encéfalo, se
extienden al diencéfalo, a la médula espinal y a la corteza cerebral.
El Sistema reticular ocupa el espacio que queda entre los núcleos segmentarios,
las vías ascendentes, y las vías descendentes, y va a estar constituido por los siguientes
núcleos:
• Núcleos reticulares del Bulbo.
• Núcleos Reticulares del Puente
• Núcleos Reticulares del Mesencéfalo
• Núcleos Reticulares del Diencéfalo.
Este sistema reticular es, pues, un centro de convergencia de las vías de
información del hombre que se envían desde todo el SNC. Las conexiones entre el
sistema reticular y las diversas partes del SNC se establecen a través de dos tipos de
neuronas, de forma que se configuran dos tipos de vías:
a) Vías de axón largo: ascienden o descienden hacia la corteza o la médula.
b) Vías de axón corto: coordinan los núcleos segmentarios y modulan funciones
vitales.
151
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CLASIFICACIÓN DE LOS NÚCLEOS DE LA FORMACIÓN RETICULAR
DEL TE
Los núcleos del sistema reticular del tronco encefálico pueden dividirse en tres
grandes grupos, tal y como a continuación se presenta:
1. Núcleos reticulares relacionados con el cerebelo.
2. Formación reticular central.
3. Núcleos reticulares del Rafe o Medianos.
NÚCLEOS RETICULARES RELACIONADOS CON EL CEREBELO
Los núcleos reticulares relacionados con el cerebelo y sus relaciones principales
se presentan a continuación:
• Núcleo Reticular Lateral del Bulbo: se sitúa al lado de la oliva bulbar y
establece conexiones con el cerebelo.
• Núcleo Reticular Paramediano: se sitúa a nivel de la zona medial o interna
del bulbo, y también envía fibras hacia el cerebelo.
• Núcleo Reticular o Calota Póntica: se sitúa ya a nivel mesencefálico.
De esta manera, los núcleos de conexión con el cerebelo se establecen a lo largo
de todo el TE y dan lugar a un circuito retículo-cerebeloso-reticular (envían la
información al cerebelo y regresan a la formación reticular). Van a emitir también
axones largos, que parten principalmente de la calota póntica y del núcleo paramediano
NÚCLEOS RETICULARES CENTRALES
Se sitúan a lo largo de toda la porción dorsal del TE, es decir, a nivel del bulbo,
de la protuberancia y del mesencéfalo. Van a clasificarse en función de las conexiones
que establecerán:
a) Núcleos con Conexiones Largas: con médula y corteza cerebral, a través de
unos haces de fibras que son el retículo-espinal lateral (inhibidor de los
músculos extensores) y el haz retículo-espinal anterior (facilitador de los
extensores, estimula el movimiento). Los núcleos que dan conexiones largas
son:
152
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
- Núcleo Reticular Central del Bulbo.
- Núcleo Reticular Gigantocelular.
- Núcleo reticular Pontis Oralis.
- Núcleo Reticular Pontis Caudalis.
b) Núcleos con conexiones cortas: los núcleos principales que van a dar
conexiones cortas son:
- Núcleo Reticular Dorsal del Bulbo.
- Núcleo Reticular Parvocelular del Puente: modula o coordina núcleos
segmentarios del bulbo
Estos núcleos van a estar conectados a través de un haz o fascículo, que se
conoce como Fascículo Central de la Calota, situado a ambos lados del tronco cerebral.
NÚCLEOS RETICULARES DEL RAFE
Se sitúan también a nivel del bulbo, del puente y de la sustancia gris situada a
nivel del acueducto de Silvio. Se localizan a nivel de la zona media (los anteriores eran
dorsales). Este tipo de núcleos establece conexiones largas con la corteza y conexiones
cortas con los núcleos segmentarios. Los núcleos reticulares del rafe son:
a) Núcleo Oscuro del Rafe.
b) Núcleo Pálido del Rafe.
c) Núcleo Magnus del Rafe.
d) Núcleo Pontino del Rafe.
e) Núcleo Central Superior del Rafe.
f) Núcleo Dorsal del Rafe.
FASCÍCULOS DE LA FORMACIÓN RETICULAR
Uno de los haces de la formación reticular es el Retículo-Espinal establece
relaciones con la médula espinal. No obstante, existen dos fascículos de dicho haz
retículo-espinal:
• El fascículo lateral es inhibidor, pues inhibe los extensores actuando a nivel
del circuito gamma de la médula. Así, si existe una afección de este haz, se
produce hipertonía muscular.
153
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• El fascículo anterior es excitador, por lo que si se produce lesión a este nivel,
se originará hipotonía.
El segundo de los fascículos que configuran la formación reticular es el haz
Rubro-Espinal, que procede el núcleo rojo e inhibe la vía piramidal. Finalmente, nos
encontramos con el haz Vestíbulo-Espinal, que establece conexiones desde el cerebelo y
el TE.
FUNCIONES DEL SISTEMA RETICULAR
Las principales funciones que va a llevar a cabo la formación reticular son las
que presentamos a continuación:
• Control del circuito gamma: actúa con el fascículo retículo-espinal lateral
como inhibidor y con el fascículo retículo-espinal anterior como
estimulador.
• Correlación en cerebelo: a través de conexiones vestibulares.
• Estimulación de la corteza: mediante conexiones colinérgicas.
• Interviene en el comportamiento motivado.
• Interviene en el reflejo de orientación y de investigación: activa el sistema de
alerta del organismo.
• Interviene en la selección de la información.
• Contiene centros vegetativos vitales.
ESTADOS DE COMA
El conocimiento de la formación reticular permite la valoración clínica de los
estados de consciencia, ya que éstas pueden ser producidas de diversas formas:
• Pérdida de la consciencia: disfunción en parte superior de protuberancia o
mesencéfalo.
• Disfunción de pares craneales: al encontrarse éstos en sitios específicos, en
los núcleos segmentarios, la exploración neurológica puede indicarnos el
nivel de la lesión en el TE.
De esta manera, es posible comprender los dos tipos de estados de coma (o
ausencia de consciencia). Son los siguientes:
154
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
3. Postura de rigidez de decorticación: lesión por encima del núcleo rojo, por
lo que la persona está en flexión (los reflejos de extensión son inhibidos).
4. Postura de rigidez de cerebración: lesión por debajo del mesencéfalo. Se
afecta el núcleo rojo, por lo que los reflejos extensión no son inhibidos, y por
lo tanto, el paciente aparece en posición de extensión.
SUSTANCIA BLANCA DEL TRONCO ENCEFÁLICO
El TE, como cualquier parte del sistema nervioso central, va a estar compuesto
por sustancia gris y sustancia blanca. Una vez conocida la estructura de la sustancia gris,
comenzaremos con el estudio de la materia blanca. Ésta se divide en dos grandes
grupos, que son:
1. Vías que atraviesan el TE: se trata de una serie de vías que se encuentran a lo
largo del todo el tronco encéfalo. Pasan por él, pero no se originan en el TE.
Las más importantes son:
- Vía piramidal y Geniculada: estas vías descienden por la parte
anterior de los pedúnculos cerebrales, de forma vertical por el puente
y llegan hasta la médula.
- Vía sensitiva: tactil-epicrítica y propioceptiva consciente. Ascienden
por los haces Goll y Burdach, por detrás de la piramidal, dando lugar
al Lemnisco Interno.
- Vía de la sensibilidad termoalgésica: se sitúa por detrás de la tactil-
epicrítica. En el mesencéfalo lateral va por detrás del lemnisco
interno.
- Vías espinocerebelosas: lateralmente, van a los pedúnculos
cerebelosos superior inferior y medio.
2. Vías propias: se originan en el propio tronco encéfalo y se dirigen hacia
regiones diversas. Las más importantes son:
- Cintilla longitudinal Posterior: une todos los núcleos que tienen que ver
con los movimientos oculares y que están relacionados con la médula
espinal, subtálamo y tronco encéfalo. Va a estar relacionada, por lo tanto,
con la coordinación de movimientos oculares en consonancia con los
movimientos de cabeza y cuello, así como con el sistema vestibular,
coordinando los movimientos de la audición.
155
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
- El haz de la calota (compuesto por dos semihaces): une los núcleos
reticulares.
- Cintilla Longitudinal Dorsal o de Schultz: es la más posterior y está en
íntimo contacto con las cavidades (en la parte más posterior, a la altura
del suelo del cuarto ventrículo y el acueducto de Silvio). Une todos los
centros vegetativos del TE e hipotálamo.
TOPOQUIMIA DEL TRONCO ENCÉFALO
Es la distribución topográfica de neurotransmisores y neuromoduladores en el
TE. En el TE existen centros con distinta topoquimia neuronal y de vías nerviosas que
transportan sus neurotransmisores a lugares más o menos alejados, bien por un
transporte y difusión por vía sanguínea o por el LCR. Existen receptores para estas
sustancias en diversidad de órganos. Pueden actuar como neurotransmisores o como
neuromoduladores (actúan sobre otras partes del SNC que producen otros
neurotransmisores)
GRUPOS DE SISTEMAS NEUROQUÍMICOS
Los principales sistemas neuroquímicos son los que enumeramos a continuación,
seguidos de su localización en el tronco del encéfalo:
• Sistema noradrenérgico (A1-A7)
• Sistema adrenérgico (C1-C3)
• Sistema dopaminérgico (A8-A15)
• Sistema serotoninérgico (B1-B9)
• Sistema colinérgico (Ch1-Ch6)
• Otros centros:
- Histaminérgico.
- GABA, Glutamato, Aspartato, Glicina Taurina.
- Sustancia P, VIP, colecistoquinina, Gastrina.
- Neurotensina, Neuropéptidos Hipofisiotropos.
- Derivados de la Pro-Opiomelanocortina, Encefalinas.
156
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Ahora que ya conocemos la morfología externa del cerebelo, es hora de
comprender como es su constitución interna. Así, cuando realizamos un corte de
cerebelo se observa cómo la sustancia blanca se sitúa en el centro, formando el famoso
Árbol de la Vida, y la sustancia gris está formando unas laminillas, y aparece en la
corteza y en los núcleos grises centrales.
En cuanto a la constitución propia de la sustancia gris de la corteza del cerebelo,
es preciso decir que se va a organizar en cinco tipos de células fundamentales. La (1)
Célula de purkinje es el tipo célula principal del cerebelo, con forma de pera y que
posee un árbol dendrítrico característico. Todas ellas se sitúan en un plano y poseen
muchas espinas dendríticas (presentan gran número de sinapsis). Además se
caracterizan por poseer un axón que parte de la corteza del cerebelo y que se suelen
dirigir a los núcleos centrales. Todas las órdenes que salen del cerebelo lo hacen a partir
de las células de purkinje. Además, podemos añadir que los cuerpos de estas células se
van a situar de forma ordenada en una sóla capa, que conforma ya de las tres que
forman el cerebelo, que son:
1. Capa de las células de purkinje.
2. Capa molecular: existen gran cantidad de fibras y relativamente pocas células.
En la capa molecular, además de las dendritas de las células de purkinje,
existen otros dos tipos de células distintas. Uno de ellos son las (2) Células
estrelladas externas (o células de Cajal) y el otro son las (3) Células
estrelladas internas (o células en cesta). Lo característico de las células en
cesta es que poseen un axón que se sitúa siempre pegado al cuello de las
células de purkinje, formando una especie de “cesto” (de ahí el nombre de las
células).
3. Capa de los granos: en esta capa existen muchas células pequeñas, de ahí el
nombre de la capa. Presenta dos tipos de células principales: por un lado está
la (4) Célula de Golgi y por el otro los (5) Granos del cerebelo. Estas últimas
son muy pequeñas (sobre 7 micras) y tiene un axón que se dirige hacia arriba,
hacia la corteza, donde se divide en dos ramas formando una T y establece
sinapsis con células de purkinje. Estos axones se sitúan perpendicularmente al
plano de las células de purkinje, de forma que recuerdan a los hilos de un poste
de luz. Son, pues, fibras paralelas a la superficie del cerebelo, de ahí su
157
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
nombre de Fibras Paralelas. Estas células tienen un sistema sináptico
especial, que es el glomérulo cerebeloso. Al glomérulo cerebeloso llegan unas
fibras que son las Fibras
Musgosas que llegan desde
fuera del cerebelo y se
introducen en él (desde la
oliva bulbar), y que hacen
sinapsis con las células de
los granos. No obstante, hoy
se sabe que este glomérulo
posee una cápsula, y que
existe algo más que una
simple sinapsis, pues en él
aparecen también axones de células de Golgi, o incluso dendritas de las células
de Golgi.
Una vez conocidas las células constitutivas del cerebelo, cabe decir que a él van
a llegar dos tipos de fibras:
a) Fibras musgosas: que proceden de la oliva bulbar y que, cuando pasan próximas
a núcleos centrales, suelen enviar colaterales.
b) Fibras trepadoras: ascienden y se distribuyen como una “hiedra” alrededor de las
células de purkinje, con la que establece sinapsis. También envía colaterales a
núcleos centrales.
158
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CONEXIONES BÁSICAS DEL ARQUICEREBELO
El arquicerebelo es el cerebelo vestibular o del equilibrio, ya que sus conexiones
se establecen precisamente con los núcleos vestibulares y el sistema del equilibrio.
La mantención del equilibrio en el organismo humano se lleva a cabo mediante la
acción conjunta de tres sistemas, que son:
• Cerebelo y Núcleos Vestibulares
• Sensibilidad Propioceptiva.
• Vista.
El arquicerebelo recibe información directa de los núcleos vestibulares y la
procesa. Las Células de purkinje contactan con el núcleo del techo, y éste a su vez, se
conectará con los núcleos vestibulares. Éstos establecerán sinapsis con los pares
craneales o con la médula a partir del fascículo vestíbulo-espinal.
CONEXIONES BÁSICAS DEL PALEOCEREBELO
El paleocerebelo, en cuanto a sus conexiones, debe asociarse con el cerebelo
espinal, ya que recibe información propioceptiva incosciente (de tendones, músculos,
articulaciones….) a través de los fascículos espino-cerebeloso ventral y dorsal. El
fascículo ventral entra en el cerebelo por el pedúnculo cerebeloso superior, mientras que
el dorsal lo hace por el pedúnculo cerebeloso inferior. Existe un tercer fascículo
denominado “cúneo-cerebeloso” que también penetra en dicho emplazamiento. Los tres
fascículos se dirigen a la corteza del paleocerebelo.
La corteza del cerebelo contacta con el núcleo globoso y el emboliforme (los dos
del paleocerebelo), a través de los cuales la información llega al núcleo rojo, la
formación reticular e incluso a la oliva cerebelosa. Estos núcleos enviarán también a la
médula ciertas órdenes para controlar el sistema motor. El núcleo rojo inhibe la
extensión de la extremidad superior y facilita la flexión. En la formación reticular, se
encargan de controlar los movimientos gravitatorios. Por ello, el cerebelo se encarga de
mantener posiciones en contra de la gravedad. De esta manera, el cerebelo es capaz de
coordinar el movimiento muscular en relación con el tono muscular y la postura.
159
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CONEXIONES BÁSICAS DEL NEOCEREBELO
Éste podría denominarse como cerebelo cortical, pues sus funciones y
conexiones están en relación con la corteza cerebral motora, ya que existe un circuito
que comienza en la corteza cerebral, pasa por el cerebelo, y termina en la corteza
cerebral de nuevo. Esto tiene que ver con el movimiento voluntario y el aprendizaje
motor.
De la corteza, el circuito desciende hasta el puente, mas concretamente hasta sus
núcleos propios. Los núcleos del puente envían unos axones (cruzados) a la corteza del
neocerebelo a través del pedúnculo cerebeloso medio. La información llegará a la
corteza cerebelosa y luego a la oliva cerebelosa, que enviará fibras al tálamo para que
la información llegue de nuevo a la corteza motora. Este sistema suele enviar ramas
colaterales, algunas de ellas hacia el núcleo rojo. La mayor parte de las fibras son
cruzadas.
SÍNDROME CEREBELOSO
Ahora qua ya sabemos las funciones y los circuitos anatómico-funcionales
básicos del cerebelo, podemos deducir cuáles serán los efectos de una lesión cerebelosa
determinada en un paciente:
a) Si tiene afectado el arquicerebelo: el paciente tiene trastornos del equilibrio,
como marcha atáxica (se observa en el individuo al caminar,…), que se
caracteriza porque se aumenta el plano de sustentación y la persona anda a
tumbos: “marcha del borracho”. Esta marcha atáxica también se puede
aumentar al cerrar los ojos. Aunque los síntomas son parecidos a las lesiones
de los cordones posteriores de la médula, se pueden distinguir los enfermos
atáxicos de los que tienen afectados los cordones posteriores. Así, en el
síndrome de Romberg, como no existe sensibilidad propioceptiva, el
paciente se cae. El enfermo cerebeloso se mueve mucho más, pero no llega a
caerse, ya que el sistema propioceptivo aún lo conserva.
b) Nistagnus: son movimientos rápidos de los movimientos oculares (pueden
ser horizontales, verticales…) Puede haber nistagmus de origen cerebeloso,
pero lo común es que sean de origen vestibular.
c) Hipotonía: es típica de una lesión cerebelosa.
160
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
d) Trastornos den la coordinación del movimiento voluntario:
- Dismetría: no mide bien los movimientos voluntarios (no son capaces de
atinar con un objeto o tocarse la punta de la nariz).
- Asinergia: este término hace referencia a que todos los movimientos
poseen un tiempo, pues actúan varios músculos para lo que es necesario
que se contraigan y relajen de forma ordenada y adecuada, con las
intensidades oportunas. Si falla el sistema, no hay sinergia sobre los
músculos, pues aunque se contraen igual, no lo hacen de forma correcta
(se produce entonces la asinergia). Esto se estudia mediante movimientos
coordinados (mandamos al paciente hacer prono-supinación o flexión-
extensión de algún miembro).
- Adiadococinesia: la sinergia del cerebelo actúa en todos los movimientos
del cerebelo. Cuando la sinergia se estudia en movimientos contrariados,
la falta de sinergia recibe el nombre de adiadococinesia.
- Temblor intencional: se produce un temblor cuando se intenta realizar
un movimiento voluntario. No se da en reposo. Esto se produce porque
no es capaz de coordinar todos los músculos (flexores y extensores) ni en
tiempo ni espacio.
- Disartria cerebelosa
- Pérdida de aprendizaje motor: se estudia con una diana y gafas de
distorsión.
Cuando se compaginan la dismetría y la asinergia, el paciente no es capaz, por
ejemplo, de parar un movimiento
determinado a tiempo. Igualmente, le cuesta
frenar una vez que empieza a caminar. Uno
de los problemas de la sinergia y la dismetría
es el habla. Hablan de forma muy rara, pues
no poseen coordinación de la musculatura del
habla (pronuncian más lentamente,
vocalizando mal).
161
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El tálamo óptico es un núcleo ovoide situado por debajo del trígono a ambos
lados del tercer ventrículo y por debajo de la prolongación frontal del ventrículo lateral;
y a su lado presenta una estructura de materia gris conocida como Sustancia Enrejada de
Arnaud.
El tálamo óptico va a estar formado por una lámina de sustancia blanca, la
Lámina Medular Interna. Por debajo, presentará otra lámina que lo separa de la
sustancia reticular y que es la Lámina Medular Externa. Finalmente, en la parte interna
(en sus caras ventriculares), presenta una pequeña lámina de sustancia blanca que recibe
el nombre de Estratum Zonale. Así, se forma la Cápsula Talámica, que recubre al
tálamo (formada, pues, por la lámina medular externa y el estratum zonale).
Existen numerosas clasificaciones para los núcleos de los tálamos. Nosotros
emplearemos una clasificación que se rige por criterios clínicos, debido a su mayor
utilidad en el campo de la medicina.
La lámina Medular Interna, se divide en la zona anterior en “Y” y llega a la
superficie. En la zona posterior, también tiene lugar esta división en “Y”, pero en este
caso los brazos de esa Y no alcanzan la superficie. De esta forma, la lámina medular
interna nos va a dividir el tálamo en varios grupos nucleares:
162
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
- Núcleo anterior: interviene en el circuito límbico o de Papez, que parte del
tálamo, pasa por el Gyrus Cinguli y llega al hipocampo. Del hipocampo,
llega a los tubérculos mamilares y, finalmente, desde aquí se conecta con el
tálamo mediante el fascículo mamilo-talámico. Al lesionar este sistema se
produce falta de memoria y conducta animal.
Entre la lámina medular externa y la interna no existirá únicamente un núcleo,
sino que se encuentran varios grupos de núcleos. Por un lado, encontramos un grupo
que se sitúa por encima y que llamaremos Núcleo dorso-lateral, y por otro
distinguiremos un núcleo situado debajo del primero, que llamaremos Núcleo ventro-
lateral. Cada uno de estos núcleos presentan una serie de características definitorias:
- En el núcleo ventro-lateral se observa la existencia de otros subnúcleos (tres
concretamente):
→ Ventro-lateral anterior (situado en la parte anterior): este núcleo
está conectado con el sistema de los núcleos caudado y lenticular, y
también en parte con la sustancia negra. Por ello, forma parte de los
circuitos que tienen que ver con el movimiento asociado al movimiento
voluntario.
→ Ventro-lateral lateral (situado en la parte media): tiene relación con
los circuitos del cerebelo (específicamente con el circuito
neocerebeloso).
→ Ventro-lateral posterior (situado en la parte posterior): Se trata de
un núcleo que posee gran importancia, ya que es el núcleo sensitivo
por excelencia. Hay que recordar que todas las vías de la sensibilidad
consciente tienen una unidad talámica (salvo la olfatoria). Este núcleo
ventrolateral posterior presenta en su parte interna una zona especial,
que se dedica a la sensibilidad de la cara, una zona que se conoce como
núcleo semilunar o arcuato, y en él terminan las fibras sensitivas del
trigémino. Es lo que se conoce como lemnisco trigeminal.
- El Núcleo dorso-lateral: es miotalámico, y está en relación con el desarrollo
de la corteza cerebral. Recibe información de los núcleos ventro-laterales y
la dirige directamente hacia la corteza cerebral.
Por dentro de la lámina medular interna y hasta el estratum zonale existe una
estructura que se conoce como Núcleo Medial. Se trata de un núcleo que interviene en
los procesos entre hipotálamo y corteza.
163
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Por dentro del estratum zonale, entre el epitelio ependimario y el estratum,
existen regueros celulares que se conocen como Núcleos de la Zona Media, que en el
hombre están muy pocos desarrollados. Se parecen a los núcleos del hipotálamo, por lo
que muchos autores consideran que estos núcleos están conectados con el hipotálamo.
Debemos señalar también que a Nivel posterior existen tres núcleos a nivel
posterior: el pulvinar, en la parte superior, y los cuerpos geniculados, en la inferior.
- Núcleo pulvinar: es un núcleo hipotalámico, sobre todo relacionado con la
corteza parietal. Según la teoría del profesor Delmás, este núcleo nos permite
conocer nuestro esquema personal (saber que somos nosotros y no otros). Para
otros estudiosos, sin embargo, el núcleo pulvinar tiene relación con los
movimientos oculares (como considera el Profesor Acuña).
- Cuerpos geniculados lateral y medial: están en relación con dos grandes vías: la
auditiva y la óptica. Se trata de estaciones de empalme de dichas vías. El cuerpo
geniculado lateral está en relación con la visión, mientras que el cuerpo
geniculado medial está en relación con la audición.
VÍAS AUDITIVAS
En este apartado se pretende especificar cual es el sistema de transmisión de los
impulsos sensitivos procedentes de la audición desde el oído interno hasta el cerebro.
Así, el procedimiento sería el siguiente:
164
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El órgano de Corti recibe información, y se lleva a los núcleos cocleares, en la
región póntica del suelo del cuarto ventrículo. Desde aquí se envía la información a la
corteza cerebral, concretamente a las áreas auditivas. Estas áreas auditivas recogen la
información, pero no van directamente a la corteza, sino que pasan por el tálamo óptico
(por el cuerpo geniculado medial). Las fibras que conectan el tálamo con las áreas
auditivas de la corteza se denominan Radiaciones Auditivas de Fayfer. La vía presenta
estaciones intermedias que no serán comentadas, ya que presentan una importancia
mínima para el médico, pues los oídos envían información mezclada a ambos
hemisferios, por lo que no se puede realizar un diagnóstico clínico de donde se
encuentra una lesión de las vías auditivas.
Únicamente existe algo diferencial entre lo que reciben los hemisferios, pero se
centra más bien en el procesamiento. El hemisferio izquierdo interpreta mejor las
palabras, mientras que el hemisferio derecho interpreta mejor aquellos aspectos
relacionados con la entonación de las palabras. No obstante cabe decir que la
información llega a ambos hemisferios, lo que pasa es que cada uno de ellos se
especializa en un procesamiento distinto.
SUSTANCIA ENREJADA O EMPARRILLADA DE ARNAUD Y OTROS NÚCLEOS
Del tálamo parten una serie de fibras que llegarán a todas las zonas de la corteza,
de forma que la sustancia gris del tálamo queda formando como “Rejas”. Esto conforma
la Sustancia Enrejada de Arnaud.
Del mismo modo, existen unos núcleos de células situadas en la lámina interna,
uno de ellos más importante que el resto, y que en conjunto forman los Núcleos
Interlaminares.
Ambos núcleos forman parte de lo que se conoce como sistema reticular
talámico, cuya función es la comunicación con la corteza cerebral y activación de la
misma. Forma parte, por tanto, del Circuito de Activación Ascendente.
165
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
LESIÓN DEL TÁLAMO ÓPTICO
Dada la gran variabilidad de núcleos y funciones que presenta el tálamo óptico,
en función de la situación de la lesión nos vamos a encontrar con patologías
tremendamente variadas:
- Lesión de los cuerpos geniculados: el paciente presentará lesiones de la vía
óptica o la auditiva.
- Alteraciones del núcleo anterior: alteraciones de la conducta y en ocasiones de la
memoria.
- Lesión del núcleo ventrolateral: alteraciones en los movimientos
No obstante, y pese a toda esta variabilidad de patologías, hemos de recordar al tálamo
en relación con la Sensibilidad, pues es el centro subcortical más importante de la
misma. En relación con esto, podemos tener dos problemas que son contrarios:
- Así, si se lesiona el núcleo ventrolateral posterior se pierde toda la sensibilidad
menos la auditiva, la visual y la olfatoria.
- Existe un filtro del dolor en el tálamo (además del que existe en la médula).
Cuando hay lesión en el tálamo, es posible que tenga lugar una Hiperpatía de
Fernández. Los sujetos que la padecen pueden presentar pérdida de la
sensibilidad y, al mismo tiempo, sufrir dolor. Es lo que se conoce como Dolor
Central. Así, cualquier estímulo sensitivo produce dolor. Forma parte de lo que
se conoce como Sindrome Talámico. Esta alodinia se cree que es provocada
porque hay fibras de la corteza cerebral que descienden y que establecen un
sistema similar al de la puerta del dolor medular. Si esta vía se lesiona es lo que
produce (muy probablemente) el síndrome talámico.
166
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En el hombre, la corteza es la más desarrollada de todos los animales terrestres,
y gracias a ella tenemos las capacidades cerebrales superiores (habla, previsión,
juicios,…) Existe una norma clásica (que no es verdad al 100%) que dice que todo lo
consciente en el hombre debe llegar a la corteza cerebral. Esto es un axioma clásico,
pues hoy día sabemos que podemos, por ejemplo, sentir dolor gracias a regiones
hipotalámicas. El lóbulo que más se ha desarrollado en el hombre es el frontal (que
supone el 40% de toda la superficie).
Para su estudio, hemos de tener en cuenta que cuando hablamos de corteza nos
referimos a sustancia gris, que presenta un grosor variable (de 2 mm a 5 mm). Esa
sustancia gris que es la corteza cerebral va a estar estructurada en varias partes, que
presentarán funciones diferentes. Como ya sabemos, existen muchas clasificaciones
para la corteza, pero la más común es la división por áreas. Este tipo de clasificaciones
suelen regirse por criterios histológicos. El sistema más empleado en clínica es el de
Brodman, que considera que existen 52 áreas, que se diferencian en las neuronas que las
componen. No obstante hay zonas de transición entre unas áreas y otras.
TIPOS DE CORTEZA CEREBRAL
ALLOCORTEX
El allocórtex o corteza primitiva es mínimo en el hombre, y está compuesta por
dos tipos de corteza:
a) Archicórtex: es un córtex compuesto por tres tipos de células:
a. Células granulares: son células estrelladas que tienen la función
principal de recibir información (tienen muchas espinas y sinapsis).
Serán abundantes en áreas sensitivas.
b. Células piramidales: serán regiones efectoras, que mandarán
información y órdenes a otros lugares.
c. Células polimorfas: fusiformes, Células de Martinoti (en toda la
corteza, estrelladas y con axón largo),…
b) Paleocórtex.
167
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
ISOCÓRTEX
El isocórtex es neocorteza que estará compuesta por seis capas diferenciadas,
que son las siguientes (se nombran de afuera a dentro):
1. Capa molecular: compuesta en su gran mayoría por fibras, aunque presenta
algunas células de Cajal.
2. Capa granulosa externa: recibe también el nombre de capa de las células
grandes o gigantopiramidal.
3. Capas piramidal externa: también se denomina capa de las células pequeñas.
4. Capa granulosa interna.
5. Capa piramidal interna.
6. Capa de las células fusiformes o capa polimórfica: células con dos penachos
y un gran axón.
Esta gran cantidad de capas es el resultado de la existencia de comunicaciones
entre distintas regiones. Así, la granulosa interna recibe información de las grandes vías
del cuerpo, mientras que la vía piramidal interna va a enviar órdenes (motoras u otras) a
otras grandes zonas del cuerpo. Las capas externas, por el contrario, son las que tienen
relación con otras áreas de la corteza. Así las piramidales dan órdenes a otras partes de
la corteza cerebral (aunque a veces llegan al hipotálamo).
TIPOS DE ISOCÓRTEX
Atendiendo a la proporción de las células que lo componen, existen dos tipos de
isocórtex diferentes:
168
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
a) Isocórtex Homotípico: seis capas celulares proporcionadas.
b) Isocórtex Heterotípico: seis capas con predominio de un tipo celular
- Córtex granular o koniocórtex (De polvo) - áreas 1, 2 y 3: predominio
casi absoluto de las capas granulosas. En áreas sensitivas.
- Córtex piramidal o agranular: predominio muy alto de las células
piramidales.
- Córtex parietal: se trata de corte de asociación, fundamentalmente.
El isocórtex homotípico va a predominar en zonas cuya función no va a ser ni
sensitiva ni motora, sino que tendrán funciones de asociación (áreas psíquicas, de
reconocimiento,…). Es muy frecuente, pues, a nivel parietal y frontal. El córtex
heterotípico granular será abundante en las regiones sensitivas, mientras que el córtex
heterotípico piramidal va a presentarse en regiones motoras.
MESOCÓRTEX
Se trata de una zona mixta con regiones de allocórtex y de isocórtex. Nos lo
encontraremos en el Parahipocampo y el Gyrus Cinguli.
DISTRIBUCIÓN DE LAS FIBRAS EN LA CORTEZA CEREBRAL
Algunas de las fibras están distribuidas de forma perpendicular a las capas. Son
las fibras radiadas, a las que se les da mucha importancia hoy día. Esto se debe a que
existe otra estructuración de la corteza, pues sabemos que el cerebro, además de estar
ordenado en “líneas”, lo estará también en “columnas”, lo que se conoce como
Columnas de Montcastle. Las columnas respondían a una misma sensación o a un
mismo estímulo. El número de células en sentido vertical es siempre la misma, salvo en
las áreas visuales (que es el doble que en otras áreas). De esta forma, para los fisiólogos,
la corteza cerebral esta organizada en columnas. Así, aunque histológicamente no se
observa una estructura, si funcionalmente (columnas de Montcastle).
Igualmente, existen una serie de fibras horizontales que reciben el nombre de
estrías. Son las siguientes:
a) Estría Superficial.
b) Estría de Kaes-Betcherev (situada en la tercera capa).
169
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
c) Estría de Baillager externa (en la cuarta capa): está muy desarrollada a nivel
del área visual (área 17), donde se puede observar mediante un corte
histológico.
d) Estría de Baillager interna (en la quinta capa).
Las neuronas de proyección son las que van a partes alejadas del cuerpo
(aferentes, eferentes y asociaciones corticales comisurales - cuerpo calloso, trígono).
Las demás neuronas son de asociación entre las dos “partes” de la corteza (externa e
interna).
AREAS DE BRODMAN
La clasificación de brodman es la más utilizada en los estudios médicos y en los
procedimientos clínicos, por lo que será la que especifiquemos. Dicha clasificación
establece 52 áreas cerebrales, entre las cuales cabe destacar:
→ Area 4: se trata de la región motora primaria.
→ Áreas 6 y 8: áreas promotoras, en las que se encuentran los programas
motores de nuestros movimientos. Están situadas por delante del área 4.
→ Áreas 9, 10, 11: es la región pre-frontal, y es un área de gran importancia en
los fenómenos psíquicos: idea personal de las cosas, juicio, capacidad de
prever proyectos vitales y demás,… Aquí actúan todos los sistemas que
pretenden influir sobre el psiquismo humano (educación,…). Como
consecuencia de ello, hace años, a los enfermos psíquicos se les extirpaba el
lóbulo frontal (lobotomía). Hubo un paciente al que le salto una piedra al
lóbulo frontal, por lo que los frenos sociales emocionales desaparecieron (no
le tenía cariño a los hijos, dejó a la familia, hacía todo lo que le gustaba,…).
170
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
→ Áreas 44,45: son áreas en las que se encuentran los centros del habla. Es el
Área De Broca. Por encima de dicha área, se sitúa un centro que presenta
todos los esquemas motores mediante los cuáles podemos escribir. Cuando
estos programas fallan, aparece la agrafia: los pacientes no son incapaces de
escribir. Hay personas que poseen dobles centros (normalmente se tienen en
un solo hemisferio, el contrario al que se escribe). Esto nos lleva a hablar de
la lateralización. Nosotros tenemos dos hemisferios cerebrales, y casi todos
los puntos de los mismos están relacionados punto a punto. No obstante,
existen sistemas, como es el caso de los motores, que precisan de unas
órdenes. Estas órdenes las da sólo un hemisferio. Por ello, el hemisferio que
da las órdenes es el Hemisferio Dominante Motor. Existen aspectos
lateralizados (habla, escritura,…) y otros que no lo están.
→ Áreas 3,1,2: al área tres llega la información sensitiva y las áreas uno y dos
reconocen cómo son las cosas.
→ Área 5 y 7: reconoce las cosas. Es el área de las Gnosias. Dentro de las
gnosias está el reconocimiento facial. Estas zonas, con el área 8 y 6, están en
relación con las praxias (movimientos automatizados habituales). Un fallo
en esto, produce apraxias.
→ Área 39 y 22: unen los conceptos con las palabras. Cuando falla este
mecanismo, aparece lo que se conoce como afasia sensitiva o de Wernich.
No nos permite hablar con normalidad, pues se dicen palabras que no tienen
sentido. No obstante si son capaces de entender. Cuando se lesiona sólo el
área 39 se produce la ceguera verbal. Son capaces de ver la palabra, pero no
entender la escritura.
→ Área 17: es el área estriada o de la visión.
→ Área 21: área de la fisión.
→ Área 18: periestriada.
→ Área 19: paraestriada.
→ Las áreas del gyrus cínguli están en relación con paleocéfalo.
→ Áreas 38 y 28: áreas de la olfacción.
Como apunte importante, hemos de señalar que las áreas de Wernich y Broca
están unidas por el fascículo longitudinal superior.
171
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CUERPO CALLOSO Y ASIMETRÍA CEREBRAL
En cinco pacientes a los que se le había cortado el cuerpo calloso, se realizó un
estudio que consistía en comunicarse con cada uno de los hemisferios del individuo
por separado (se le ponían imágenes en un ojo, sin que lo viera el otro). Así, se les
pedía a los hemisferios que realizaran funciones diferentes. De esta manera, cuando
conectamos con el hemisferio dominante, se elaborará una respuesta, mientras que si
va al otro, el individuo no responderá. De esta manera, se llevó a cabo un estudio
sobre la lateralización de las funciones cerebrales:
HEMISFERIO IZQUIERDO HEMISFERIO DERECHO
Líneas Rectas y Escritura Dibujo artístico
Audición para el habla Audición para la entonación y la música
Lenguaje y habla
Cálculo Percepción espacial
172
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Finalmente, sólo señalar que nuestro sistema nervioso funciona como un sistema
digital y analógico al mismo tiempo. El hemisferio derecho compara cosas
(analógico), mientras que el hemisferio izquierdo funciona como un ordenador
digital (mueve cifras, números, letras,…). Por ello todos tenemos una parte
“artística” y una parte “práctica”; pero ambos funcionan conjuntamente con el
cuerpo calloso, por eso respondemos como una persona única. No obstante sabemos
que cada hemisferio podría funcionar de forma independiente.
173
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Para comenzar con el estudio del oído, diremos ya desde ahora que su función
es transducir las ondas sonoras y transformarlas en impulsos nerviosos. Las ondas
sonoras tienen características: la frecuencia (ciclos/s) y la intensidad (decibelios). De
esta manera, podemos decir que los sonidos cuya frecuencia se encuentre entre los 50 y
los 16.000 Hz serán audibles por un individuo joven. Con la edad, suele aparecer
presbiacusia, que es la pérdida de captación de las frecuencias más altas.
El oído humano está mucho más adaptado para frecuentas entre 300 y 3.000
ciclos por segundo, es decir, el intervalo en el que se incluyen los sonidos producidos
por el aparato fonador humano.
La necesidad de la existencia de dos oídos proviene de la necesidad de saber de
dónde proceden los sonidos, ya que éstos llegarán antes a un oído que al otro.
En cuanto al desarrollo de los oídos, hemos de decir que provienen de los órganos de la
línea lateral, que son como “bolsas” de líquido que captan las vibraciones que se
transmiten por el agua. En el proceso de evolución, estos órganos de la línea lateral
tuvieron que adaptarse a la vida terrestre, en donde los sonidos se transmiten por el aire
en lugar de por el agua; ya que el aire presenta una capacidad de transmisión del sonido
mucho menor.
En lo que se refiere a la estructura del oído humano, cabe decir que debemos
distinguir tres partes bien diferenciadas con funciones características. Estas tres partes
son:
• Oído Externo: capta las ondas, las recoge y las dirige hacia la zona interna.
Presenta, pues, un efecto de antena.
• Oído Medio: cavidad que presenta una cadena de huesecillos que transmite
las ondas sonoras al oído interno. Por ello recibe el nombre de transmisor.
• Oído Interno: lugar en el que se produce la transducción de las vibraciones
a impulsos nerviosos.
Haciendo una referencia general a la patología, podemos decir que si no existiera
el oído medio, la audición seguiría siendo posible, pero se perdería gran capacidad
auditiva. En relación con este aspecto, podemos decir que existen dos tipos
fundamentales de sordera que se diferencian en la región del oído afectada:
1. Sordera causada por patología del oído medio: no se transmiten bien las
vibraciones, por lo que se produce una sordera de transmisión.
174
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
2. Sordera causada por patología del oído interno: en este caso la sordera es
de origen neurológico y se produce por un funcionamiento neuronal
incorrecto o ineficaz.
EL OÍDO Y EL EQUILIBRIO
Los órganos de la línea lateral que antes hemos comentado eran también
utilizados para darse cuenta de la posición corporal y mantener el equilibrio. En el ser
humano, esta función persiste en el oído, pero se ha especializado, originándose el
Sistema Vestibular, encargado de los dos tipos de equilibrio (estático y dinámico).
EL OÍDO EXTERNO
El oído externo va a estar compuesto por dos estructuras bien diferenciadas que
son las que presentamos a continuación:
1. Oreja: es el elemento más externo del oído.
2. Conducto auditivo externo: finaliza al llegar al tímpano.
LA OREJA
La oreja va a estar compuesta, a su vez, por una serie de estructuras bien
diferenciadas que son las que a continuación se enumeran y explican:
• Pabellón auditivo: presenta forma ovoide, y se presenta formando un ángulo
con la cabeza, que se conoce con
el nombre de ángulo céfalo-
auricular y que no debe ser mayor
de 30º.
• Hélix: canal que forma el reborde
de la oreja. Tiene forma cóncava.
• Antihélix: se trata de una
estructura convexa que deja una
depresión conocida como fosilla
escafoidea.
175
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
• Trago: es una prominencia que presenta, en su cara interna, un mechón de
pelos cuya función es la protección del oído, y que recibe el nombre de
Bárbula Hirci.
• Antitrago: se encuentra situado en una posición inferior con respecto al
trago.
• Concha: es la cavidad más grande de la oreja.
• Escotadura de la concha o trágica: situada entre el trago y el antitrago.
• Lóbulo: emplazamiento especialmente indicado para extracciones de sangre.
La oreja tiene una capa de piel por fuera y cartílago elástico en el interior. LA
piel presenta muy poco tejido celular subcutáneo. Cabe decir también que en la cara
externa existen fibras que unen directamente la piel con el cartílago, de ahí que los
procesos en esta cara de la oreja puedan llegar a ser muy dolorosos.
Como un simple detalle, añadiremos que, aunque existen músculos que
permitirían mover el pabellón auditivo, en el hombre no están desarrollados, por lo que
la mayor parte de la población no es capaz de mover las orejas.
En las observaciones del oído con un otoscopio, es necesario tirar de la oreja
hacia arriba y hacia atrás, para introducir el mismo; así como echar el trago hacia
delante.
CONDUCTO AUDITIVO EXTERNO
Se trata de un conducto muy irregular, no rectilíneo, que presenta una curva
convexa hacia arriba longitudinalmente. En el plano transversal, el conducto auditivo
externo se dirige hacia delante, hacia atrás después y finalmente de nuevo hacia delante.
El conducto auditivo externo presenta una serie de relaciones muy importantes
desde el punto de vista clínico, ya que algunas de ellas pueden ser causa de dolor de
oído. Así tenemos las siguientes relaciones:
• Cara superior: está en relación con la fosa craneal media.
• Cara parotídea o inferior: está en estrecha relación con la glándula parótida.
Por ello, en patologías de la parótida, el paciente suele sufrir dolor de oídos.
• Cara anterior: relacionada con la articulación temporomandibular. Los
dolores de la ATM también pueden simular dolor de oídos.
• Cara posterior: en relación con la apófisis mastoides.
176
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En cuanto a la estructura del conducto auditivo externo, hemos de decir que en él
se diferencian dos partes:
1. Externa: es fibrocartilaginosa, y representa un tercio del volumen total del
conducto.
2. Interna: es ósea, y configura los dos tercios internos del conducto.
La parte ósea presenta una parte superior formada por la porción escamosa del
temporal, que se conoce con el nombre de Segmento de Rivinus. El resto de la
superficie es el Hueso Timpanal o Timpánico.
En lo que se refiere a la parte fibrocartilaginosa, cabe decir que está estructurada
en dos capas, una en la parte inferior formada por cartílago; y otra en la parte superior,
formada por tejido fibroso. Hay que especificar que en la parte cartilaginosa existen
unas estructuras que reciben el nombre de Tendiduras de Duvernay. Igualmente, va a
presentar una capa de piel en su interior con glándulas sebáceas y sudoríparas. Algunas
de estas últimas forman un sudor muy espeso que cuando se seca forma el Cerumen.
Esta sustancia forma unos “tapones” que pueden ser eliminados con agua a presión
(este chorro de agua debe ser dirigido hacia la cara superior)
La inervación del conducto es complicada, ya que la zona de Ramsey-Hunt está
inervada por el glosofaríngeo (IX par) y el pneumogástrico (X par). Por ello, cuando se
estimula el vago (con agua fría, por ejemplo), se reduce la frecuencia cardíaca y la
presión arterial. Este suceso provoca mareos.
177
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
EL OÍDO MEDIO
Es la parte del oído que se sitúa entre el oído externo y el interno, y está
compuesto por tres partes:
1. La caja del tímpano.
2. Las celdas mastoideas: amplían las vibraciones.
3. Comunicación con la faringe mediante la Trompa de Eustaquio: la trompa o
tuba se encarga de comunicar la faringe con el oído, haciendo que la presión
a un lado y al otro del tímpano sea la misma.
LA CAJA DEL TÍMPANO
Se trata de una cavidad situada oblicuamente, y que se sitúa entre el oído externo
y el interno. Contiene una cadena de huesecillos relacionados con dos músculos muy
importantes para la audición, que son el Músculo del Martillo y el Músculo del Estribo.
Está tapizada por una mucosa y contiene vasos y nervios.
La caja presenta una forma aplanada de fuera adentro (su altura es de un
centímetro y medio), de tal forma que, en la zona más estrecha, tiene muy pocos
milímetros y se va ensanchando. Los consideraremos, no obstante, como un cubo
aplanado, de forma que, para su estudio, tendremos en cuenta seis paredes: superior,
inferior, externa, interna, anterior y posterior. Estas caras van a recibir denominaciones
específicas.
PARED EXTERNA O TIMPÁNICA
Da al conducto auditivo externo, y en ella va situada la membrana timpánica. La
Membrana Timpánica está rodeada por un marco óseo que está compuesto por la
escama del temporal (alveolo de los huesecillos) en la parte superior, y por el hueso
timpánico en el resto de la superficie. La parte de la escama del temporal que forma el
techo del conducto auditivo externo se denomina Segmento de Rivinus. Entre ellos
aparece un surco en el que se fija la membrana del tímpano.
La membrana se inserta básicamente en el hueso timpánico, y casi nada en la
escama. Igualmente, en su interior se observa una parte de un hueso, el Martillo, con su
cabeza, cuello, apófisis corta y apófisis larga (que termina en una zona ensanchada que
178
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
es la Espátula del martillo). La membrana del tímpano se fija fuertemente al hueso
martillo, así como al hueso timpánico. Esta última fijación se lleva a cabo mediante
fibras conjuntivas que se mezclan con fibras conjuntivas del hueso, formando un rodete
fibroso que recibe el nombre de “Rodete anular de Gerlach”.
Desde el martillo salen dos ligamentos, el Ligamento Tímpano-maleolar
Anterior y el Ligamento Tímpano-maleolar Posterior, que llegan a la altura de la
escama. Por debajo de los ligamentos se sitúa la Pars Tensa, porque siempre se
encuentra en tensión y está agarrada al martillo. La superficie que queda por encima de
los ligamentos se conoce como Pars Flácida.
Cabe decir, no obstante, que la membrana timpánica posee tres capas: una
primera que es del epitelio que recubre el conducto auditivo externo, una interna que es
de mucosa y la media, que está compuesta por fibras
conjuntivas (la propia membrana). Sin embargo, a la parte
superior le falta la capa conjuntiva, de ahí que reciba el nombre
de Pars Flácida (sólo tiene mucosa y epitelio). Esta parte forma
una especie de “bolsa” que recibe el nombre de Bolsa Superior
de Prussac.
La Pars Tensa se mantiene en su estado característico de tensión gracias a la
existencia de fibras elásticas y colágenas (estas últimas en mayoría) que lo permiten.
Aparecen de esta forma unas fibras radiadas y fibras redondeadas en toda la superficie
de la membrana. Igualmente existen otros dos tipos de fibras, que son las fibras
parabólicas y las fibras semilunares. Las primeras se inician en el mango del martillo y
llegan al rodete del Gerlach del lado contrario; y las segundas van de una parte del
rodete de Gerlach hacia otra parte del propio rodete.
La espátula del martillo coincide con la parte más interna de la membrana
timpánica, en una invaginación cónica de la misma hacia la caja del tímpano (zona del
ombligo). Por lo tanto, la membrana timpánica se une firmemente al mango del martillo,
al que transmite las vibraciones ocasionadas por el sonido.
Podemos deducir, pues, que la membrana timpánica es oblicua, y mira hacia
abajo y hacia fuera. Dicha oblicuidad, varía desde unos 20-30º en el feto hasta unos 45º
en un individuo adulto. Se realizaron estudios para comprobar si
existía relación entre la audición y la oblicuidad de la membrana, y
se llegó a la conclusión de que cuanto más vertical es la
179
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
membrana, más facilidad se tiene para la audición.
Desde el punto de vista médico, la membrana timpánica es muy importante, ya
que es la primera región que se explora cuando existe algún problema del oído. Cuando
se realiza una exploración por el otoscopio, se observa que el color de la membrana se
aproxima al gris perla, un tono amarillento; además de apreciar algunos de los
elementos del oído medio gracias a que es muy fina y traslúcida. De esta manera,
cuando empleamos el otoscopio observamos el martillo (la apófisis externa, el mango y
la espátula), así como los ligamentos tímpano-maleolares. Bajo la espátula, se distingue
un cono de luz, conocido como Cono Luminoso de Wild, y que sólo se observa en los
tímpanos sanos, ya que en un oído patológico, la luz del otoscopio no se refleja
correctamente. A veces puede reconocerse la arteria del mango del martillo. La
membrana del tímpano es muy fina y translúcida (mide sólo una décima de milímetro),
por lo que en ocasiones podemos observar alguna estructura del oído medio.
Siguiendo con la clínica de la membrana del tímpano, cabe especificar que para
la realización de una Paracentesis de la Membrana (apertura de la misma para llegar a la
caja del tímpano) se debe acceder a través de los cuadrantes III y IV, ya que por I y II
pasan vasos y nervios. Estos cuadrantes se establecen gracias al trazado imaginario de
dos líneas, una que pasa por el mango del martillo y otra por el ombligo. El acceso por
los cuadrantes III y IV facilita también el drenaje, ya que en ellos la fuerza
gravitacional es mayor.
CARA INTERNA O LABERÍNTICA
La cara interna del oído medio está en relación con el oído interno, una zona que
se conoce como Laberinto.
En primer lugar, observamos en la parte anterior e inferior una zona que protuye
un poco hacia fuera y que recibe el nombre de Promontorio. Dicho promontorio es el
resultado de la existencia del caracol óseo justo en dicha zona y presenta un pequeño
Orificio en la parte inferior por el que sale del oído medio en Nervio de Jacobson (el
nervio sensitivo principal de la caja del tímpano). En ocasiones (no siempre), del
orificio puede partir un surco que se divide en seis, por donde pasan las seis ramas que
da el Nervio de Jacobson justo a la salida del agujero.
Por encima del promontorio, nos encontramos con otra zona que protuye,
producida por el Conducto del Músculo del Martillo. Dicho conducto, en su parte más
180
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
posterior, presenta un orificio que se conoce con el nombre de Proceso Cocleariforme (o
pico de cuchara), por el que el músculo martillo accede a la caja del tímpano.
En la zona más alta, aparece la prominencia de otro conducto, y que se conoce
con el nombre de Acueducto o Nerviducto de Fallopio, en el interior del cual va el
nervio facial. En el 15% de los casos (como media), este nerviducto de Fallopio tiene
fisuras (microscópicas o macroscópicas) y no es completo. Estas fisuras comunican el
nerviducto con la caja del tímpano, algo de importancia clínica si consideramos que el
nervio facial puede infectarse a partir de afecciones de la caja del tímpano,
ocasionándose parálisis. Para corregir dichas parálisis, en ocasiones es necesario
descomprimir el nervio.
Debajo del nerviducto nos encontramos con una depresión que se conoce con el
nombre de Fosa Oval, y dentro de ella un agujero que tiene forma de platina de horno y
que se conoce como Agujero Oval. En condiciones normales este hueso está cerrado por
el estribo y por el ligamento anular de la platina del estribo.
Debajo existe otra depresión redondeada, que es la Fosa Redonda, con un
orificio en su interior que se conoce con el nombre de Ventana Redonda. Esta ventana
está cerrada por una membrana que recibe el nombre de Tímpano Secundario.
Entre las dos fosas, existe una zona que se conoce con el nombre de Seno
timpánico, que está separado de la ventana oval y de la ventana redonda por dos puentes
óseos.
Por último, en la parte más alta, por encima del nerviducto de Fallopio, existe
otra prominencia que es el resultado del conducto semicircular externo.
CARA ANTERIOR O TUBOCAROTÍDEA
En ella cabe resaltar dos aspectos: por un lado su relación con la carótida
interna, que se mete en el peñasco del temporal y sale por el vértice; y por otro, que en
ella se encuentra el orificio de la Trompa de Eustaquio o tuba faringo-timpánica. De
estos dos aspectos nace la denominación de la cara anterior de la caja del tímpano como
cara tubocarotídea.
En dicha cara, en un primer lugar, nos encontramos con el Orificio de la Trompa
de Eustaquio. Para continuar, podremos distinguir también con un Orificio para el
Nervio de la Cuerda del Tímpano (orificio anterior). Se trata de un nervio que atraviesa
la caja timpánica de lado a lado cuya función es fundamentalmente gustativa (aunque
181
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
también lleva alguna fibra vegetativa). Finalmente, existe un Filete Simpático que parte
de la carótida y que pasa por uno o dos agujeros que se conocen con el nombre de
Agujero/s Carótido-timpánico/s.
CARA POSTERIOR O MASTOIDEA
Se relaciona con todas las celdas mastoideas. En esta cara nos encontramos con
un orificio de forma triangular que se conoce como Orificio del Additus ad Antrum, que
da a un “corredor” o “pasillo” que conecta la caja del tímpano con el Antro Mastoideo,
que a su vez conectará con las Cavidades o Celdas Mastoideas (cuya función es
ampliar las vibraciones). Las celdas mastoideas se clasifican según su posición en
relación con dicho antro mastoideo. Algunas de las celdas mastoideas están próximas al
facial o incluso tocando el nerviducto de Fallopio, por lo que cuando se opera en las
celdas mastoideas es preciso tener en cuenta la relación de dichas estructuras con el
facial. Por ello se monitoriza el nervio facial en intervenciones quirúrgicas.
Igualmente se observa una prominencia ósea en forma de cono que se conoce
con el nombre de Pirámide, y que está agujereada. Presenta un conducto por el que pasa
un músculo, que es el músculo del estribo.
Existe también la Prominencia o Eminencia cordal, ya que en su centro se
encuentra el orificio posterior de la cuerda del tímpano. Finalmente la apófisis estiloides
marca una pequeña tuberosidad en la parte inferior de la cara.
Entre la eminencia cordal y la pirámide existe una cresta (Cresta Cordal), y una
segunda entre la tuberosidad ocasionada por la estiloides y la pirámide (Cresta
Piramidal). Entre ambas crestas se distingue una depresión conocida como Fosilla de
Grivault .
CARA SUPERIOR O TEGMEN TÍMPANI
Es la más craneal, y se encuentra en relación con la fosa craneal media (que da
alojamiento al lóbulo esfenotemporal del cerebro). De esta cara hemos de saber que se
conoce como Tegmen Tímpani, y en ella existe una estructura que es la sutura petro-
escamosa interna, que desaparece en el adulto (entre la porción petrosa y la escamosa
del temporal).
182
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CARA INFERIOR O YUGULAR
Su relación más importante es la que se establece con el golfo de la vena yugular
interna, que marca una prominencia en la cara inferior. Se trata de una cara irregular con
muchas espículas óseas.
INTERIOR DE LA CAJA DEL TÍMPANO
El interior de la caja del tímpano posee numerosas estructuras, entre las que se
encuentran vasos y nervios, la cadena de huesecillos, los músculos y tendones que
mueven los huesecillos, y finalmente la mucosa de la caja.
CADENA DE HUESECILLOS
La cadena de huesecillos del oído está compuesta por tres pequeños huesos que
son los que a continuación se enumeran, y que posteriormente se explican:
• Martillo.
• Yunque.
• Estribo.
183
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El Martillo.
El martillo es un hueso que está compuesto por una cabeza, un cuello y un
mango (que se fija la membrana timpánica) con una dilatación que se conoce con el
nombre de espátula. Presenta dos apófisis: una corta que se dirige hacia fuera en la
membrana del tímpano, en la que se insertan los ligamentos tímpano-maleolares, y otra
larga que se dirige hacia delante.
Yunque.
Este hueso se compara a una muela con dos raíces. Está compuesto por el cuello,
con una cabeza cóncava donde se aloja el martillo. Igualmente, presenta una apófisis
horizontal que se dirige hacia atrás y se introduce en la escotadura que hay en la entrada
del Additus ad Antrum, una fosa que recibe el nombre de Incudis. Finalmente está la
apófisis lenticular, mediante la cual se articula con el estribo.
Estribo.
El estribo tiene una cabeza esférica que se articula con el yunque y de la que
parten dos ramas: una anterior y otra posterior. Estas dos ramas se relacionan con una
parte plana, que es la platina del estribo y que se introduce en la fosa oval.
Estos tres huesecillos que acabamos de nombrar están fijos por ligamentos, tal y
como se expresa ahora:
• Ligamento Súpero-externo: se encarga del agarre del martillo
• Ligamento Posterior: sostiene el yunque
• Ligamento de la Platina del estribo.
Los huesos se articulan entre sí mediante artrodias, de forma que el yunque se
articula con la cabeza del martillo, y la apófisis lenticular del yunque con el estribo.
Estas articulaciones, que en un principio poseen cápsula articular, acaban por
desaparecer con el tiempo, formándose así una Palanca Osicular. Las funciones de la
palanca osicular es la transmisión de la vibración de la membrana timpánica hacia los
líquidos del oído interno, más concretamente a la perilinfa.
Existen dos músculos que regulan los movimientos y vibraciones de esta cadena
osicular, que son el Músculo del Martillo y el Músculo del Estribo. El músculo del
martillo se dirige hacia fuera (sale de la cara interna por el pico de cuchara) y tira hacia
adentro, de forma que tensa la membrana timpánica, y hace que la presión en los
184
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
líquidos laberínticos aumente cuando tira de los huesecillos. El músculo del estribo, por
el contrario, sale hacia abajo, de forma que saca el
estribo de los líquidos laberínticos y reduce la presión.
Igualmente hace disminuir la tensión en la membrana
timpánica, ya que tira hacia adentro. Por lo que
acabamos de explicar, se deduce fácilmente que el
músculo del martillo y el músculo del estribo son
antagónicos, aunque Los músculos funcionan siempre, coordinadamente. De hecho,
cuando hay poco ruido, o se contraen muy poco los dos, o se relaja el músculo del
estribo.
Un aspecto importante es que, ante ruidos fuertes, los músculos tiran
fuertemente para evitar grandes vibraciones. Esta potente contracción puede producir
una hipoacusia, dado que estos músculos pueden afectarse.
En los sonidos agudos, aunque se contraen los dos músculos, lo hace sobre todo
el músculo del martillo. Por el contrario, el del estribo permite escuchar sonidos graves.
Los músculos son muy pequeños, y aunque la fuerza muscular es proporcional a
su sección, lo es a su sección funcional y no a la anatómica. Son músculos penniformes,
pues aunque son pequeños, hay que sumar el calibre de las fibras oblicuas a lo largo de
todo el trayecto del músculo.
Estos músculos están inervados por el facial (en el caso del músculo del estribo)
y por la rama mandibular del V par (en el caso del martillo).
Finalmente, cabe decir que en el interior de la caja, además de los huesecillos y los
músculos, nos vamos a encontrarnos con otros elementos, como es el caso del Nervio de
Jacobson o la Cuerda del Tímpano. Igualmente, podemos nombrar la Mucosa que
recubre la caja timpánica.
CELDAS MASTOIDEAS Y TROMPA FARINGO-TIMPÁNICA
La función principal de la trompa es comunicar el exterior con la caja timpánica,
de forma que permita su aireamiento. Además, existen glándulas en el interior de la
cápsula y demás que generan unos detritus que deben ser excretados por la tuba
timpánica (por eso posee cilios). Por ello, también realiza la función de drenaje la caja.
Finalmente, en la pared de la tuba (cerca de la faringe sobre todo), existe gran cantidad
185
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
de ganglios linfáticos, lo que se conoce como Amígdala Linfática o Tubárica de
Gerlach. Son los nódulos linfoides que existen en la trompa de Eustaquio.
La trompa presenta forma de diábolo, formada por un cono óseo y un cono
fibrocartilaginoso. El primero ocupa el tercio póstero-externo de la misma y el
cartilaginoso los dos internos. En cuanto a la región de Fibrocartílago es de interés
señalar que la sección superior y externa es cartilaginosa, mientras que el resto de la
sección es fibrosa.
Es de resaltar la existencia de dos músculos pegados a la trompa. Son los
Periestafilinos Externo (pared externa de la trompa) e Interno (pared interna de la
trompa). Cuando estos músculos se contraen permiten la aireación de la trompa, ya que
la abren. Cabe especificar, no obstante, que el periestafilino es un tensor del velo del
paladar y el periestafilino interno es un elevador del velo del paladar.
El agujero de la trompa en la faringe está a un centímetro detrás del cornete
inferior, y tiene forma triangular con tres repliegues que llevan el nombre de Salpingos,
y que separan el orificio de la trompa con el resto de estructuras faríngeas. Son:
• Salpingo nasal.
• Salpingo laríngeo.
• Salpingo estafilino.
EL OÍDO INTERNO
El oído interno, como estructura funcional, va a desarrollar dos funciones
diferenciadas, que son el Equilibrio y la Función auditiva. Ambas son funciones
totalmente diferentes, aunque existen patologías de la audición relacionadas también
con afecciones del equilibrio (dado que se encuentran en la misma localización).
En el oído interno hay un líquido denominado perilinfa, muy similar al líquido
cefalorraquídeo, a la que se transmite la vibración de la cadena de huesecillos. La
vibración de la perilinfa hace que vibren membranas del caracol, deforma que se
producirá una vibración que servirá de estímulo. La perilinfa se sitúa entre el laberinto
membranoso y el hueso.
Dentro del laberinto existe otro líquido, que es la endolinfa, cuya composición es
totalmente diferente a la perilinfa y muy similar a la de los líquidos intracelulares (posee
gran cantidad de potasio y poco sodio). La endolinfa se encuentra dentro del laberinto
membranoso.
186
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Para abordar el estudio del oído interno, vamos a considerar una serie de
cavidades y conductos en el hueso, que es el laberinto óseo, así como unos conductos
membranosos que se encuentran en su interior. De esta manera, podemos dividir el oído
interno en continente (laberinto óseo) y contenido (membranas donde se sitúan los
órganos, que es el laberinto membranoso).
LABERINTO ÓSEO
Presenta una cavidad central que se conoce con el nombre de vestíbulo óseo, del
que parten una serie de conductos que son los conductos semicirculares óseos. Son los
siguientes:
1. Conducto semicircular anterior o superior: se dirige hacia arriba
2. Conducto semicircular posterior.
3. Conducto semicircular lateral o externo: se dirige hacia la parte externa
Del vestíbulo sale también un conducto que da dos vueltas y media sobre sí
mismo y que constituye el caracol óseo. Finalmente nos encontramos con el conducto
endolinfático óseo.
EL VESTÍBULO
Tiene forma más o menos cúbica o cuboidea, a modo de saco. Cabe resaltar el
hecho de que aquí se situará la ventana oval, tapada por la platina del estribo.
A nivel del vestíbulo existen dos fosillas, una superior y otra inferior, que llevan el
nombre de fosilla semiovoidea y fosilla hemisférica. Es de interés recordarlas porque en
cada una de ellas se sitúan estructuras del laberinto membranoso. Además, a nivel de
estas fosillas, en sus paredes internas, existen una serie de pequeños orificios que se
conocen como Orificios de Mancha Cribosa, por donde llega el nervio vestibular. En la
fosilla superior se aloja el utrículo y en la inferior el sáculo. Por ello los orificios de la
superior son para el nervio utricular y los orificion de la inferior para el nervio sacular.
Estas fosas están separadas por una cresta importante, porque de la que parte el
nervio utricular. Dentro del utrículo y el sáculo existen unas calcificaciones (otolitos),
que en bipedestación sedimentan de forma distinta según la posición, permitiéndonos
localizarnos en relación con la gravedad.
187
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CONDUCTOS SEMICIRCULARES
Presentan una zona estrecha y otra no estrecha. La no estrecha se llama
ampollar, y en ella hay gran cantidad de orificios para nervios. Cada uno de los
conductos presenta nervios propios. Cabe especificar que la zona ampollar de los
conductos superior y posterior es común.
CARACOL ÓSEO
Es un tubo que da dos vueltas y media y que está abierto. Si realizamos una sección, nos
encontramos con un eje central en el que se apoya el caracol para poder dar las dos
vueltas y media de espiral, que se denomina Columbela. El hueso que rodea a cada una
de las vueltas de espira que se conoce como Lámina de los Contornos.
Existe igualmente una lámina que se sitúa en el conducto óseo del caracol que se
conoce como Lámina Espiral. De esta manera, el caracol queda dividido por la lámina
espiral. Se presenta como una lámina abierta en el medio que deja un conducto
conocido como Conducto espiral de Rosenthal. Dicho conducto es tremendamente
importante, pues en él se sitúa un ganglio nervioso. Se trata de un ganglio especial que
se sitúa a lo largo de todo el conducto y que se conoce como Ganglio Espiral de Corti.
En este ganglio se sitúa la primera neurona de las vías auditivas y recibe información
del Órgano de Corti, que enviará hacia el SNC a través de los orificios de la columbela.
Los axones de las neuronas que se sitúan en el ganglio de Corti salen y se unirán para
formar un gran nervio, que es el VIII par. De esta manera, la base de la columbela es
una zona cribosa que adopta una forma espiral. Así se forma la Criba Espiroidea del
caracol (la base de la columbela va al conducto auditivo interno).
En el caracol óseo existe un pequeño orificio que comunica con el interior del
cráneo que se conoce con el nombre de acueducto del caracol (parte inferior de la
imagen).
CONDUCTO ENDOLINFÁTICO ÓSEO
Se llama acueducto del vestíbulo y recibe este nombre porque aloja en su interior
al conducto endolinfático.
188
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
EL LABERINTO MEMBRANOSO
El laberinto membranoso va a estar constituido por el vestíbulo membranoso
(sáculo, utrículo y conducto endolinfático), por los conductos semicirculares
membranosos y por el caracol membranoso. Todas estas estructuras están conectadas
entre sí de forma que:
• Los tres Conductos Semicirculares membranosos llegan al utrículo.
• El Utrículo se conecta con el Sáculo a través del Conducto Endolinfático
(este tiene dos ramas que se juntan para formar el conducto), que llega a la
fosa subarcuata. Algunos autores definen una válvula de Bass para controlar
presiones entre ambas estructuras (no está demostrado).
• El sáculo conecta con el caracol mediante el Conducto Reuniens de Hensen.
CONDUCTOS SEMICIRCULARES MEMBRANOSOS
Se sitúan en las tres direcciones del espacio y son perpendiculares entre sí.
Presentan una zona dilatada (ampollar) y una más estrecha. En la zona dilatada aparecen
los receptores del equilibrio. Son estructuras en las que aparecen células con cilios en un
magma gelatinoso formando una cresta auditiva que se moverá en función de la
localización de la cabeza a causa de los líquidos.
189
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CARACOL MEMBRANOSO
Situado en el interior del caracol óseo, donde ocupa el espacio situado entre la
lámina espiral ósea y la lámina de los contornos, completando así la separación entre la
rampa vestibular y la rampa timpánica. Se origina en la cavidad vestibular en un fondo
de saco, en donde a través el canal reuniens comunica con el sáculo, se introduce en el
orificio vestibular de la cóclea ósea y la recorre en sus dos vueltas y media de espira. En
el interior de caracol óseo existen tres conductos o rampas, la Rampa Timpánica y la
Rampa Vestibular (que contienen
perilinfa) y comunican entre ellas a nivel
del Helicotrema (cúpula o vértice del
caracol óseo) y el caracol membranoso o
Rampa Coclear (que contiene endolinfa).
El caracol membranoso adopta al
corte una forma triangular, presentando
tres caras: externa, superior e inferior.
1. Cara Externa
Se halla en contacto con la lámina de los contornos, cuyo periostio se engruesa
para formar el ligamento espiral. En el ligamento espiral se halla una zona muy
vascularizada, la estría vascular, cuya función es la formación de la endolinfa.
2. La Cara Superior.
La cara superior del caracol membranoso está formada por la membrana
vestibular o Membrana de Reissner, que es una lámina conjuntiva tapizada por un
endotelio. Se inserta medialmente en la lámina espiral, y lateralmente en el ligamento
espiral.
3. Cara Inferior
Está formada por la membrana espiral basilar, que también se inserta en la
lámina espiral y en el ligamento espiral. ES sobre la membrana espiral basilar en donde
se asienta el órgano de Corti, órgano nervioso receptor de los sonidos.
190
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Funcionalmente, las vibraciones transmitidas a través de la platina del estribo
llegan a la perilinfa, éstas vibraciones de la perilinfa permiten la vibración de la
membrana basilar y deforman el conducto coclear, es esta vibración y deformación la
que estimula las células auditivas del órgano de Corti.
La membrana basilar no tiene la misma anchura en todo el recorrido del caracol,
siendo más ancha en la zona del vértice y más estrecha en la zona de la base. La anchura
de la membrana basilar en la base del caracol es de 0,04 milímetros, y en el vértice es de
0,5 mm. Esta diferencia de anchura de la membrana basilar a lo largo del caracol
membranoso es uno de los factores que permite la discriminación auditiva.
191
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
El sentido de la visión va a estar desempeñado por un órgano que es el globo
ocular. Existe una capa nerviosa externa que es la retina, donde se encuentran los
transductores ópticos. No son más que células nerviosas encargadas de transformar las
señales nerviosas en impulsos nerviosos que serán transportados a través de las vías
ópticas hasta el lóbulo occipital del cerebro, lugar donde se encuentran las áreas
visuales.
El globo ocular va a tener también un sistema de protección que va a estar
compuesto por los párpados y la secreción de la glándula lacrimal, y un aparato motor,
formado por los seis músculos, que nos va a permitir movilizarlo en todas las
direcciones.
Tiene un peso entre siete u ocho gramos, se encuentra en la cavidad orbitaria y
se va a mantener en posición porque va a estar sujeto por unos ligamentos, por la propia
musculatura extraocular y sobre todo por la presencia de la grasa orbitaria. Existen tres
compartimentos grasos: uno entre la musculatura y otros a los lados.
Puede ocurrir que en enfermedades o patologías en las que aumenta la cantidad
de grasa, el ojo puede aparecer como salido hacia fuera (ojos saltones), lo que en clínica
se denomina exoftalmia (por ejemplo en el
hipotiroidismo). Por el contrario, en patologías
en que disminuye la grasa, el ojo aparecerá
hundido en la órbita, lo que se conoce con el
nombre de endoftalmia (es por ejemplo, un
signo de deshidratación, por lo que es
importante en lactantes).
Otra característica del globo ocular es su consistencia. Esta consistencia es muy
firme en el vivo, y va a estar producida por los líquidos que contiene en su interior,
sobre todo el humor vítreo, que es lo que se denomina como presión intraocular. La
presión intraocular normal oscila entre 12 y 22 milímetros de mercurio, y se mide con
un aparato denominado Tonómetro. Algo que si hay que decir de la presión intraocular
es que nada tiene que ver con la presión arterial, dado que el globo ocular está formado
por tres capas concéntricas y la vascularización se sitúa en el medio. Cuando la presión
aumenta puede dañar las células de la retina y da lugar a un Glaucoma.
192
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En cuanto a la forma, cabe decir que se trata de una esfera ligeramente aplanada
de arriba abajo, y su porción anterior, denominada córnea transparente, sobresale en
relación con el resto de la superficie del ojo. Esta disposición va a implicar una
desigualdad más o menos manifiesta pero constante, de los tres diámetros del globo
ocular. Así el diámetro transversal mide 23 milímetros y medio, el vertical mide 23
milímetros y el diámetro ántero-posterior mide unos 25 milímetros. La alteración de
estos diámetros da lugar a distintas patologías. Así, por ejemplo, una alteración del
diámetro antero-posterior: si dicho diámetro está aumentado, las imágenes completas se
forman por delante de la retina, dando una patología que se conoce como miopía axil.
Si el diámetro antero-posterior está acortado, las imágenes netas se forman por detrás de
la retina, algo que en clínica se conoce como hipermetropía axil.
De esta manera, observamos que la función del globo es captar las imágenes y
trasladarlas, a través de las vías ópticas, hasta el Sistema Nervioso Central. Para realizar
está función es esencial que las lentes situadas en su región anterior enfoquen
perfectamente. Para ello, el ojo posee un poder dióptrico que es constante (excepto en el
caso del cristalino, que puede variar). Para que esas lentes
puedan enfocar, las curvaturas deben ser perfectas, de forma que
los rayos incidentes penetren sin problemas. Cuando esto no es
así, los rayos pueden desviarse, dando lugar al astigmatismo
congénito.
193
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
En el globo ocular se distinguen dos polos, uno anterior que se corresponde con
la córnea transparente, y uno posterior, que se corresponde con un punto diametralmente
opuesto un poco por encima de la salida del globo.
El círculo máximo perpendicular al eje antero posterior se denomina ecuador, y
divide al globo en dos hemisferios. Los meridianos son los círculos máximos que pasan
ala vez por los dos polos.
Algo a tener en cuenta, es que existe LCR alrededor del nervio óptico, además de
cubiertas meníngeas. Con dichas envueltas, el nervio óptio llega sin problemas hasta el
polo posterior del ojo.
EL GLOBO
En el globo ocular se van a poder distinguir dos tipos de componentes, que son
los siguientes:
a) Los que conforman paredes, capas o túnicas.
b) El contenido, lo que se sitúa en el interior de las capas.
Las capas, de fuera adentro, son tres (principales), tal y como a continuación se
presentan:
1. Capa esclero-corneal (externa y fibrosa): esta capa va a estar compuesta, a su
vez por:
- Córnea
- Esclerótica
2. Úvea o coroides (capa intermedia o musculovascular): esta capa media estará
compuesta, de atrás hacia delante:
- Coroides propiamente dicha (de ahí que la capa media en conjunto se
tienda a denominar úvea)
- Zona ciliar.
- Iris
3. Retina (capa interna, nerviosa o neural).
En el interior del globo vamos a tener una serie de líquidos y una lente, que
forman los medios transparentes y refringentes del globo ocular. De adelante atrás son:
1. El humor acuoso.
2. El cristalino.
3. El humor vítreo.
194
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
CAPA EXTERNA
Como hemos dicho ya, la capa externa o fibrosa va a estar compuesta por la
córnea y la esclera. Ahora estudiaremos cada una de estas capas por separado.
CÓRNEA
La luz se va a introducir en el globo ocular a través de la córnea, que es la
estructura ocular con una capacidad de refracción más importante, y que hace que la luz
quede enfocada sobre la retina. La córnea va a ser, a la vez, una membrana resistente y
un medio transparente. Por ello, se define como una membrana fibrosa que está
enclavada o situada en la abertura anterior de la esclerótica, con la que se va a continuar,
a nivel del limbo esclero-corneal, de la que va a diferir en su estructura solamente por la
disposición de sus fibras. Va a poseer dos caras:
- Una cara anterior: en relación con el medio ambiente, los párpados y la
secreción de la glándula lacrimal.
- Una cara posterior: que va a estar bañada por el humor acuoso, junto al que
forma una lente cuyo poder dióptrico es de aproximadamente 48 dioptrías, y
es la lente que va a concentrar o que más concentra los rayos que penetran en
el interior del ojo. Esta cara, además, va a formar el límite anterior de la
cámara anterior del globo ocular.
195
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La córnea no puede tener vasos en su interior, y se va a nutrir del oxígeno del
medio ambiente, de la secreción lacrimal y del humor acuoso. La inervación sensitiva
va a proceder del nervio nasal y de los nervios ciliares largos, pues se forma un plexo
nervioso muy consistente.
La córnea está formada por cinco capas, en una estructura “En capas de cebolla”
(tal y como lo designan los histólogos). Presenta un epitelio estratificado. Cuando hay
lesiones, no se forman cicatrices, pues no hay colágeno. Sin embargo, cuando la lesión
afecta a la membrana de Bowman, que si posee colágeno, si se pueden producir
cicatrices y perder parte de visión.
Finalmente viene la membrana de
Descemet, que cede cuando la córnea recibe
un golpe para que ésta no se lesione mucho.
Aparece también un endotelio que bombea
iones, de forma que libera líquido de la
córnea, y cuando esto falla (con la edad),
pueden producirse edemas corneales.
ESCLERÓTICA
Forma, junto con la córnea, la cubierta fibrosa del globo, y recubre las 5/6 partes
del globo. Posee un color blanco-azulado en el niño, debido a la transparencia de vasos,
en el adulto es translúcida o blanca, y en el anciano adelgaza, y por transparencia de
grasa presenta un color blanco-amarillento. En la parte interna de la esclerótica anterior
existe un pequeño saliente que es el Espolón Escleral, en el que se inserta el músculo
ciliar o músculo de la acomodación.
Estructuralmente, la esclerótica está constituida por tejido conjuntivo, y a
diferencia de la estructura corneal, sus fibras se entremezclan en todas las direcciones,
ya que en ella se produce la inserción de los músculos
extraoculares. De esta manera forman unos “ovillos” que la
hacen mucho más resistente. Así, salvo en la zona donde se
continúa con la córnea, las fibras de la esclerótica son muy
consistentes.
Como es la capa externa, todo lo que entra o sale del globo ocular debe atravesar
la esclerótica. Por ello, esta capa presenta una serie de aberturas.
196
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
- En primer lugar, tiene una abertura anterior en la que se sitúa la córnea.
Alrededor de este orificio existe un pequeño número de orificios a través de
los cuales van a penetrar los vasos ciliares anteriores.
- En su parte posterior presenta también un orificio que no puede ser hueco,
dado que la presión provocaría su desgarro. Por ello, está parcialmente
cerrado por una estructura de tejido conjuntivo que va a constituir la lámina
cribosa, y que se asemeja a una red. A través de los orificios de la lámina van
a pasar las fibras que conforman el nervio óptico, así como la vena y la
arteria central de la retina. Alrededor de este orificio se sitúan entre quince y
veinte pequeños orificios, a través de los cuales pasan las arterias y los
nervios filiares. Los nervios pueden ser largos (procedentes del nervio nasal)
y cortos (procedentes del ganglio ciliar oftálmico).
- Un poco por detrás del ecuador, existen dos orificios inferiores y dos
superiores, por los que saldrán del globo ocular unos confluentes venosos de
la coroides que son las venas o vasas vorticosas. A ellas llega toda la sangre
del globo excepto la de la retina.
LA ÚVEA
Se caracteriza por actuar como elemento nutritivo para todo el globo ocular,
excepto para la retina, que posee su propia vascularización. Va a proporcionar calor, y
posee una musculatura propia. Finalmente, también diremos que es la encargada de la
formación de líquidos, como es el caso del humor acuoso, por lo que indirectamente, es
la responsable de la presión intraocular.
Se distinguen tres partes o porciones en la úvea:
- Coroides propiamente dicha: se extiende desde la salida del nervio óptico
hasta un poco por delante del ecuador del globo ocular, donde una línea
festoneada o en dientes de sierra que se denomina Ora Serrata, marca su
límite anterior.
- Zona Ciliar: se extiende desde la ora serrata, hasta el limbo esclerocorneal.
- Iris: que es un diafragma muscular situado a nivel del limbo esclerocorneal,
y que deja un orificio central que es la pupila.
197
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
COROIDES
Ocupa los 3/4 posteriores del globo ocular, y posee una coloración pardo-rojiza
debido a que es principalmente vascular, y a ella se debe la coloración rojiza que se
observa en el sujeto vivo al examinar el fondo de ojo mediante el examen
oftalmoscópico. Funcionalmente, va a actuar como membrana nutricia del ojo, y como
una pantalla ante la luz (evita que ésta se disperse).
En algunos animales de vida nocturna, existe otra capa que se llama tapiz o
pigmentum, que está llena de pigmentos, lo que hace que en la oscuridad, cuando se le
enfoca una luz, se les observe cierto color (los gatos en verde).
ZONA CILIAR
La zona ciliar se extiende hasta la ora serrata, donde se continúa con la coroides.
Va a presentar formar triangular y estará formada por un estroma vascular, músculo liso
y un epitelio de cobertura.
La zona ciliar presenta en su zona interna unas eminencias que son los procesos
ciliares, cuya función es la producción del humor acuoso. Son un número aproximado
de 80 y miden entre 2 y 3 mm. Se encuentran separados entre sí por surcos dispuestos
radialmente que reciben el nombre de valles ciliares.
Además de los procesos ciliares, en la zona ciliar vamos a encontrar el músculo
ciliar o músculo de la acomodación que está formado por fibras musculares lisas que se
insertan por dentro en el espolón estriado. La función de este músculo es acomodar la
visión a la proximidad o la lejanía, modificando la curvatura del cristalino. Así, cuando
se contrae, afloja la tensión sobre las fibras suspensorias del cristalino, con lo que este
se desplaza hacia delante y el cristalino adopta una forma más esférica, con lo que va a
aumentar su poder de refracción (numero de dioptrías). Cabe decir que, normalmente, el
cristalino está muy aplanado debido a las fibras del músculo ciliar. No obstante, si se
vuelve más “redondo’’ se permite la visión cercana.
IRIS
El iris es un diafragma que se sitúa en el limbo esclero-corneal y presenta un
orificio central que es la pupila. Constituye la porción anterior de la úvea. Posee una
198
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
forma de disco que se encuentra perforado en su centro por un agujero circular móvil
que es la pupila cuya función es regular la cantidad de luz que penetra en el globo
ocular a través del reflejo fotomotor.
El iris va a presentar dos caras para su estudio, que son las que presentamos a
continuación:
- Cara anterior que limita por detrás a la cámara anterior. Se encuentra
dividida en dos partes, central y periférica, por una línea denominada
collarete que es siempre visible porque el iris se adelgaza a partir de ella
hacia la pupila
- Cara posterior: esta aplicada contra la cara anterior del cristalino y delimita
con este la zona ciliar y la zónula de Zinn la cámara posterior del globo
ocular.
→ La Pupila.
La pupila es un orificio circular móvil, que en condiciones normales posee un
espesor de entre 3 y 4 mm y que se adapta como un diafragma a la intensidad de la luz.
Cuando la luz es intensa y durante la visión cercana, la pupila disminuye de tamaño.
Cuando el tamaño es inferior a 1’5 mm hablamos de miosis. Por el contrario, cuando la
luz es débil y durante la visión lejana, la pupila aumenta de tamaño (es superior a 4 mm)
hablamos de midriasis.
Estructuralmente el iris estará formado por 4 capas bien diferenciadas, tal y como a
continuación se detalla:
- Membrana limitante anterior
- Estroma
- Musculo dilatador del iris
- Epitelio posterior
En el estroma, en el borde de la pupila o circunferencia menor del iris
encontramos fibras, que componen el músculo esfínter de la pupila, dispuestas en
círculo alrededor de la pupila.
La capa del músculo dilatador va a estar formada por las prolongaciones
contráctiles de células mioepiteliales presentes en su epitelio posterior. Se disponen
desde la base del iris o circunferencia mayor del iris hasta el borde pupilar, en forma
radial.
199
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Cuando se contraen las fibras circulares se produce el cierre de la pupila,
mientras que cuando se relajan las fibras circulares y se contraen las radiales se produce
la apertura de la pupila.
Un aspecto característico del iris es su color, que viene determinado por la
cantidad de melanocitos presentes en el estroma. Si hay pocos, los ojos son azules,
mientras que si hay muchos, los ojos son pardos o marrones, siendo el gris y el verde
colores intermedios. En el caso de los albinos, se cree que no presentan melanocitos( o
que si poseen las células pero por un defecto genético son incapaces de cargarlas de
pigmentos).
En la vida normal la pupila tiende a estar cerrada (miosis) debido al sistema
nervioso parasimpático, que actúa sobre ella. No obstante debemos tener en cuenta una
serie de factores que producen la contracción de la pupila, y otros que producen su
dilatación:
→ Causas de la miosis:
- Luz
- Sueño, cansancio
- Miótica (parasimpaticomimética, simpatolítica)
- Síndrome de Horner (incluido ptosis y estrechamiento de la
hendidura palpebral)
- Narcosis, morfina
- Pupila rígida
→ Causas de la midriasis:
- Oscuridad
- Excitación
VASCULARIZACIÓN DE LA ÚVEA
Como hemos dicho ya, la úvea es la capa músculo-vascular del globo ocular, por
lo tanto será importante tener en cuenta la procedencia de dicha vascularización. Cabe
comentar, por ello, que dicha vascularización va a proceder de tres orígenes:
- Arterias ciliares cortas posteriores: son entre 20 y 30. Atraviesan la
esclerótica por los orificios del nervio óptico, penetran en el interior del
globo ocular y avanzan entre la esclerótica y la coroides, donde se ramifican
en arterias coroideas. Al atravesar la esclerótica, van a suministrar algunas
200
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
ramas colaterales, las cuales, anastomosándose entre ellas, forman alrededor
del nervio óptico un anillo o circulo denominado anillo vascular escleral,
circulo de Haber o circulo arterial del nervio óptico.
- Arterias ciliares largas posteriores: son dos, una interna o nasal y una
externa o temporal, y atraviesan la esclerótica un poco por encima del
orificio que da paso al nervio óptico y avanzan hacia delante hasta alcanzar
el cuerpo ciliar, donde se dividen en dos ramas (una superior o ascendente y
otra inferior o descendente), las cuales se anastomosan 2 a 2 formando
alrededor de la circunferencia mayor del iris el circulo arterial mayor del
iris.
- Arterias ciliares anteriores: también son en número variable, suelen ser 2
para cada músculo recto, excepto para el recto externo que suele ser una. Son
ramas de las arterias musculares, perforan la esclerótica cerca de la inserción
de os músculos rectos para penetrar en el globo ocular e incorporarse al
círculo arterial mayor del iris.
Del círculo arterial mayor del iris salen numerosas ramas que se originan del
propio círculo o de las arterias que lo forman. Entre ellas destacan unas ramas que
partiendo del círculo se dirigen hacia el borde pupilar en forma radiada y al alcanzar
este se anastomosan entre ellas formando alrededor del borde pupilar el circulo arterial
menor del iris.
En cuanto a las venas, destaca el hecho de que siguen un trayecto inverso a la
red arterial y todas van a desembocar a la red coroidea, la cual drena a través de las
venas o vasas vorticosas las cuales atraviesan la esclerótica, para ir a drenar a las venas
oftálmicas.
201
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
LA RETINA
La retina es la capa mas interna de las capas del ojo. Embriológicamente es la
única que tiene origen medular y va a tener distinto grosor, pues la luz no la impresiona
por igual, por lo que su desarrollo va a ser distinto. Las zonas principales son:
- Retina sensible o porción óptica de la retina: es una zona gruesa, que ocupa
aproximadamente los 2/3 posteriores. Se extiende desde la ora serrata hasta
la papila óptica (punto por el cual las fibras del nervio óptico atraviesan la
esclerótica).
- Zona anterior o retina ciega: es la que se sitúa en el 1/3 anterior y es más
delgada. Muchos autores discuten que exista retina a este nivel.
Desde el punto de vista morfológico, la retina es una membrana fina,
transparente y ligeramente rosada en la que su cara externa se pone en contacto con la
coroides por medio del epitelio pigmentario y su cara interna se halla en contacto con el
humor vítreo, al que ese adhiere a nivel de la ora serrata y de la papila. En el resto de su
superficie existe una zona frágil y poco solida en la que la unión es mantenida
únicamente por la presión intraocular (humor vítreo), por lo que cualquier patología que
afecte, en este caso al humor vítreo, puede provocar un desprendimiento de retina. La
retina va a estar formada por células epiteliales pigmentarias, células fotorreceptoras,
células de sostén y células nerviosas. Todas ellas están agrupadas en dos capas
(clasificación de Cajal):
- Capa externa: formada por el epitelio pigmentario de la retina.
- Capa interna: la capa interna de la retina va a estar formada, a su vez, por:
1. Epitelio pigmentario.
2. Apófisis de las células fotorreceptoras.
3. Capa limitante externa.
4. Núcleos celulares de los fotorreceptores.
5. Capa plexiforme externa.
6. Capa nuclear de las células bipolares (capa
interna de Körner).
7. Capa plexiforme interna.
8. Núcleos celulares de las células ganglionares.
9. Capa de fibras nerviosas.
10. Capa limitante interna.
202
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La retina va a presentar una serie de modificaciones en áreas especializadas
específicas, que son: la mácula, la papila óptica y la retina periférica.
- Mácula: en el punto que se corresponde con el eje posterior del globo ocular
encontramos la retina central o macula lútea, la cual posee un diámetro de
entre 5 y 6 mm. En su centro va a presentar una zona de unos 3 mm
denominada fóvea central y que se observa como una pequeña depresión en
la que sólo existen conos (distinguen colores y formas).
Cada cono de dicha región hace sinapsis con una sola
neurona bipolar, y ésta, a su vez, con una única neurona
ganglionar, por lo que va a proporcionar una gran
resolución. Como esta área se encuentra en el centro
del campo visual y proporciona una visión a color muy detallada se la
denomina también punto óptico o punto de máxima visión. Otra
característica de la mácula es que presenta un color amarillento que se debe a
la presencia de pigmentos, sobre todo de los carotenoides.
- Papila óptica o punto ciego: se sitúa entre los 2’5 y 3 mm hacia el lado
interno y 1 mm por encima del polo posterior del globo ocular, y presenta
una morfología pálida y redondeada. Esta zona se corresponde al punto por
el cual los axones de las células ganglionares atraviesan la esclerótica para
dar origen al nervio óptico. Esta zona carece de fotorreceptores y por tanto
provoca un punto ciego en el eje visual. Cabe señalar
que el nervio óptico se considera una parte del SNC,
debido a que en el interior de la órbita está rodeado
por meninges, lo que quiere decir que el LCR llega
hasta el polo posterior del globo ocular. Por este
motivo, cualquier patología que curse con un aumento del LCR hace que la
papila aparezca como hinchada, lo que se denomina papila o yema (esto es
un signo físico importante que suele indicar una inflamación cerebral
patológica o traumática). La papila presenta en su centro una pequeña
depresión denominada excavación fisiológica a través de la cual los vasos de
la retina penetran en la misma.
203
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
La Vascularización de la retina va a depender de la arteria central de la retina,
que en el interior de la misma se divide en una rama ascendente y otra descendente, las
cuales a su vez se dividen en dos ramas, una interna nasal y otra externa o temporal. El
drenaje, por el contrario, se realiza a través de la vena central de la retina. Se forma a la
inversa, de forma que dos venas interna convergen hacia el centro de la papila para
formar la vena central.
En un fondo de ojo, la papila siempre se ve con unos bordes muy nítidos y
cuando hay una patología estos bordes dejan de ser tan nítidos. En la fóvea central o
punto de máxima visión, el resto de las capas se apartan y prácticamente solo queda la
parte de fotorreceptores y por tanto la luz incide casi directamente.
204
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
MEDIOS TRANSPARENTES Y REFRINGENTES DEL GLOBO OCULAR
Nos vamos a encontrar una serie de estructuras que actúan como lentes, tales
como el humor acuoso y el cristalino, y otra que recubre el globo ocular en su parte
posterior, que es el humor vítreo. En conjunto, se denominan medios transparentes y
refringentes del globo ocular.
EL HUMOR ACUOSO
Se sitúa entre el cristalino y la córnea y es el responsable principal de la presión
intraocular. Va a ocupar las denominadas cámaras del globo ocular:
- La cámara anterior, que está situada entre la cara posterior de la córnea y la
cara anterior del iris.
- La cámara posterior, situada entre la cara posterior del iris y la cara anterior
del cristalino, zona ciliar y zónula de Zinn.
Ambas capas se comunican a través del orificio pupilar, de forma que el humor
acuoso puede pasar de una cámara a otra.
El humor acuoso es un líquido similar al LCR que es secretado por los procesos
ciliares del cuerpo ciliar, a una velocidad de 2 ml/min. Desde aquí pasa a la cámara
posterior y, a través del orificio pupilar, llega hasta la cámara
anterior. Se va a reabsorber a nivel del limbo esclero-corneal
donde, por detrás del mismo, encontramos un conducto vascular
conocido como Conducto o Canal Venoso de Schlemm. Por detrás
de él, el tejido rico en colágeno de la esclera se disocia en laminillas o trabéculas,
adquiriendo un aspecto semejante a una red. Estas laminillas, en conjunto, constituyen
el Ligamento Pectíneo de Huek o Trabécullum. Las laminillas delimitan unos espacios
que son las Lagunas de Fontana, que establecen una
comunicación entre la cámara anterior y el conducto de
Gloso. De éste se van a originar unos conductos que
desembocarán en el plexo venoso epiescleral o plexo venoso
de Lebber, desde donde pasan a la circulación general.
205
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Va a existir una resistencia normal al flujo del
humor acuoso a través de la malla trabecular, por lo que la
secreción y reabsorción constantes originan una presión de
reposo normal dentro del globo ocular (que oscila entre los
10 y 22 mm de Hg). Las alteraciones en estas vías de
drenaje aumentan dicha presión y pueden originar el
Glaucoma, que si no es tratado, daña las células de la
retina (lo que conduce a la ceguera a largo plazo).
CRISTALINO
Es una lente biconvexa y transparente que se sitúa entre el iris y el vítreo y se
mantiene en posición gracias a unas fibras que lo sujetan a la zona ciliar y que en
conjunto constituyen el Ligamento Suspensorio del Cristalino
o Zónula de Zinn. La función de estas fibras, además de
mantenerlo en posición, es transmitir al cristalino la
contracción del músculo ciliar. La propiedad principal del
cristalino es la plasticidad, gracias a la cual puede modificar
su curvatura, y por tanto su índice de refracción (el número de
dioptrías), durante el proceso de la acomodación. El cristalino
va a poseer un poder dióptrico de aproximadamente 15
dioptrías y es la gran lente filtradora de rayos UV.
En el feto, es una estructura esférica, ligeramente rojiza, con una consistencia
blanda. En el adulto adquiere una consistencia más firme. Con los años el cristalino
aumenta de volumen, se hace más aplanado, adquiere un tinte amarillento y se vuelve
más duro y menos transparente. Este proceso se denomina esclerosis del cristalino, y se
caracteriza por la pérdida de agua y el aumento de su densidad, lo que lleva a una
ineptitud del cristalino para variar su forma, y por tanto su poder de refracción, durante
el proceso de la acomodación. Esto se denomina en clínica
Presbicia, Presbiopía o Vista Cansada.
Al tratarse de un órgano que no posee vasos, su nutrición
deriva de los líquidos intraoculares. Los intercambios precisos
para la nutrición del cristalino se realizan por difusión a través
206
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
de una cápsula, por lo que existe la posibilidad de que se depositen sustancias en su
interior (que ocasionaría la pérdida de su transparencia). Esta patología se conoce en
clínica como Catarata u Opacificación del Cristalino.
EL HUMOR VÍTREO
El humor o cuerpo vítreo constituye aproximadamente el 80% del volumen del
ojo (es la estructura más grande del mismo). Se trata de un gel transparente que se
localiza por detrás de la zona ciliar, el cristalino y sus zónulas; y que ocupa el espacio
limitado por la superficie interna de la retina. La característica especial del vítreo es que
se adapta a todo lo que lo rodea.
Durante el período embrionario, por el interior del vítreo discurre la arteria
hialoidea, la cual forma una red arterial por delante del cristalino. A partir del séptimo
mes, la arteria se oblitera y desaparece, permaneciendo el conducto en que se alojaba,
que recibe el nombre de conducto o Canal de Cloquet. En ciertas patologías congénitas
permanece la arteria hialoidea.
ANEXOS DEL GLOBO OCULAR
Los anexos del globo ocular se reúnen en dos grandes aparatos, teniendo en
cuenta su función. Así, tenemos los siguientes:
- El aparato locomotor: compuesto por los músculos extrínsecos, que son los que
a continuación se indican:
→ Recto superior.
→ Recto inferior.
→ Recto externo.
→ Recto interno.
→ Oblicuo Mayor.
→ Oblicuo Menor.
→ Elevador del Párpado.
- Aparato de protección: dicho aparato va a estar compuesto, a su vez, por tres
elementos diferenciados:
207
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
→ Párpados.
→ Conjuntiva.
→ Aparato lacrimal.
MUSCULATURA EXTRÍNSECA DEL GLOBO OCULAR
Se trata de un conjunto de músculos estriados que actúan a pares y que están
cubiertos por una fascia. Son el recto superior, el recto inferior,
el recto interno, el recto externo, el oblicuo mayor y el oblicuo
menor. Entre los músculos extrínsecos existe un tendón común
con dos orificios (tendón de Zinn). El orificio mayor se conoce
con el nombre de anillo de Zinn.
Estos músculos van a ser los responsables del movimiento del globo ocular, pero
hay que tener en cuenta que los diferentes movimientos oculares van a estar
determinados por una serie de características específicas, tal y como mostramos a
continuación:
• Incongruencia de los ejes ocular y
orbitario: esto va a provocar que los
músculos no tengan una acción pura y
simple. Esto no afecta a los rectos
interno y externo, que si tendrán.
• Inserciones musculares en el globo
ocular: existen movimientos que son
el resultado del tipo de inserción del
músculo que lo realiza. Por ejemplo, el
oblicuo mayor provoca una rotación y un giro (lleva el polo anterior hacia
abajo y hacia fuera), mientras que el oblicuo menor hace lo contrario.
• Existencia de unos ejes de movimiento del globo ocular: los diferentes
movimientos oculares están en relación con unos ejes:
- Eje antero-posterior: movimientos de rotación.
- Eje vertical: convergencia y divergencia.
- Eje lateral: visión a derecha o izquierda.
208
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Para comprender perfectamente la acción de los distintos músculos extrínsecos,
Márquez desarrolló un esquema que funciona cuando los ojos están en posición
anatómica, mirando al infinito; y que se basa en los movimientos que sufriría el polo
anterior del ojo cuando actuaran los distintos músculos.
Cuando existe una lesión de los nervios óculo-motores, se va a producir en el
paciente una visión doble (Diplopia), en la que los pacientes moverán la cabeza
intentando de algún modo limitar la vista a un solo campo (pues se marean en caso
contrario).
LOS PÁRPADOS
Los párpados son dos repliegues móviles que continúan los tegumentos de la
cara, y que cuando se cierran, obturan la base de la órbita y cubren la cara anterior del
globo ocular. Protegen los ojos de los traumatismos externos, los cuerpos extraños, y
contra la luz excesiva o exposición prolongada a la misma.
Los párpados están en continuo movimiento, con lo que aseguran una igual
distribución de las lágrimas y de las secreciones de glándulas, con lo que se consigue la
lubrificación del globo ocular, mantener la córnea limpia y transparente y liberar al ojo
de cualquier partícula extraña que se haya depositado en la superficie.
Cada uno de los párpados presenta una cara anterior, una posterior, un borde
libre (en que se implantan las pestañas) y un borde fijo que se continúa sin un límite
claro de demarcación con la piel de la cara y el borde inferior de las cejas.
Los bordes libres poseen una longitud de unos 30 mm y un grosor que oscila
entre los 2 y los 3 mm. Se unen hacia fuera formando el ángulo o canto externo y por
dentro formando el ángulo o canto interno. Sobre dicho borde libre, y cerca de su
209
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
ángulo interno, nos encontramos una pequeña elevación que es la Papila o Tubérculo
Lagrimal, en cuyo vértice se encuentra el Punto Lacrimal, que es donde se inician los
canalículos lacrimales superior e inferior. Estos orificios van a dividir al borde palpebral
en dos porciones:
- Una interna o lagrimal: que es corta, y se caracteriza por no tener pestañas.
Entre los tubérculos y el ángulo interno, se delimita un espacio denominado
lago lagrimal, que está ocupado por una pequeña elevación rojiza que es la
carúncula lagrimal.
- Porción externo-ciliar: es mayor que el anterior, y comprende la porción del
borde palpebral situado entre los tubérculos y el canto interno. Se caracteriza
por ser la porción en la que se implantan las pestañas.
La estructura palpebral está formada por piel, tejido celular subcutáneo, tejido
muscular, tarsos y conjuntiva. El borde libre de cada parpado presenta hacia delante un
labio redondeado del cual se originan las pestañas y cuyas raíces están provistas de
glándulas. Estas glándulas son de dos tipos:
- Glándulas de Zeis o ciliares: sebáceas
- Glándulas de Moll: sudoríparas
Cuando hay una infección aguda por invasión estafilacocica de dichas glándulas
se produce una patología conocida comúnmente con el nombre de orzuelo.
En cuanto al tejido muscular, cabe decir que en los parpados podremos
encontrarnos con dos músculos:
- Elevador del parpado: se origina en el tendón de Zinn, va por encima del
recto superior y se va a insertar en el tarso.
- Músculo orbicular: está contenido en el espesor del parpado y es un
músculo circular en el que se distinguen
dos porciones: una porción palpebral y
una porción orbital. La función es cerrar
los párpados. La inervación está
controlada por el VII par. Si hay una
patología las lagrimas caen por abajo en
lugar de pasar por delante del globo.
210
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
Los Tarsos son dos láminas alargadas de tejido fibro-elástico que le confieren a
cada párpado su firmeza. Ambos tarsos se unen por medio de dos ligamentos que los
fijan a tejido óseo y que reciben el nombre de Ligamentos Palpebrales Externo e
Interno.
En el espesor de los párpados vamos a encontrar las glándulas de Meibomio.
Estas glándulas se disponen en hileras paralelas y son unas glándulas sebáceas que
secretan una sustancia rica en lípidos que retrasa la desaparción de la película lacrimal y
además lubrica y protege la córnea. Cuando se obstruyen, no pueden secretar, y
aumentan de tamaño y se producen los quistes denominados chalacios o chalazones.
La conjuntiva recibe este nombre porque se une al globo ocular y los párpados,
presentando tres partes bien diferenciadas:
- Conjuntiva palpebral.
- Fondo de saco conjuntival.
- Conjuntiva ocular o bulbar.
APARATO LACRIMAL
En el ser humano, el aparato lacrimal se compone de dos partes diferentes: una
porción secretora, compuesta por las glándulas secretoras, y otra porción excretora, que
recoge la secreción de dichas glándulas para llevarlas al meato inferior de las fosas
nasales; y que en conjunto constituyen las vías nasales.
Las lágrimas son producidas por una glándula lagrimal principal y por las
glándulas lacrimales accesorias de Krausse (párpado) y Wolfring (conjuntiva). Así,
gracias a la secreción de dichas glándulas se forma la Película lacrimal, que no es más
211
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
lOMoARcPSD|1684066
que una fina película transparente que cubre toda la superficie externa del ojo. Todo lo
externo es temporal y todo lo interno nasal, por lo que el movimiento de los párpados
llevan las lágrimas de fuera adentro.
Las lágrimas se van a extender por la superficie de la cornea y de la conjuntiva y
se depositan en el lago lacrimal. A continuación penetran por los canalículos lacrimales
superior e inferior, que presentan dos porciones: una corta vertical y una más larga
horizontal. Ambas porciones se unen en un canal único o conducto lacrimal que va a
desembocar en el saco lacrimal. Este ultimo no es
mas que la porción superior dilatada del conducto
lácrimo-nasal, el cual va a desembocar en el
meato inferior de las fosas nasales, donde existe
una válvula denominada Plica lacrimalis o válvula
de Hassner.
212
Su distribución está prohibida | Descargado por Sofia Castillo (srodrigrezdelacruz@yahoo.com)
Potrebbero piacerti anche
- 06-TEST DEL PARCIAL N°2 INFORMÁTICA LegalDocumento15 pagine06-TEST DEL PARCIAL N°2 INFORMÁTICA Legallopezvictor01100% (1)
- 1ra, 2da y 3ra Semana de N.A.F.Documento49 pagine1ra, 2da y 3ra Semana de N.A.F.Mirel LobNessuna valutazione finora
- Tema 3 - NeuroanatomíaDocumento48 pagineTema 3 - NeuroanatomíaGissell ErazoNessuna valutazione finora
- Clase 3 Medula EspinalDocumento21 pagineClase 3 Medula Espinaltyrone_orozcoNessuna valutazione finora
- Neuroanatomía Funcional Del Sistema NerviosoDocumento55 pagineNeuroanatomía Funcional Del Sistema NerviosoEasy NeonnNessuna valutazione finora
- 6 Mesecefalo y MetencefaloDocumento16 pagine6 Mesecefalo y MetencefaloRichard Álvarez ChungNessuna valutazione finora
- MesencéfaloDocumento39 pagineMesencéfaloDianaUgaldeNessuna valutazione finora
- Wuolah Free Resumen Completo Libro Anatomia Gulag FreeDocumento27 pagineWuolah Free Resumen Completo Libro Anatomia Gulag Freepm2rn4vdm8Nessuna valutazione finora
- GUIA SISTEMA NERVIOSO CENTRAL UnisimonDocumento14 pagineGUIA SISTEMA NERVIOSO CENTRAL UnisimonCésar ObregónNessuna valutazione finora
- GUIA SISTEMA NERVIOSO CENTRAL UnisimonDocumento13 pagineGUIA SISTEMA NERVIOSO CENTRAL UnisimonCésar ObregónNessuna valutazione finora
- Apunte 2 Clase 2 Neuroanatomia y Circuitos 2023Documento7 pagineApunte 2 Clase 2 Neuroanatomia y Circuitos 2023Rafahela Antonia Vergara RiveraNessuna valutazione finora
- Reporte CerebroDocumento9 pagineReporte CerebroLizette AngelNessuna valutazione finora
- Estructura Del Sistema NerviosoDocumento15 pagineEstructura Del Sistema NerviosoSARA SOFFIA VALDIVIESONessuna valutazione finora
- GUIA DISECCION LAB BIOLOGIA ResueltaDocumento8 pagineGUIA DISECCION LAB BIOLOGIA ResueltaKaren Julieth ARIZA ORTIZNessuna valutazione finora
- Neurofisiologia Del EncefaloDocumento31 pagineNeurofisiologia Del EncefaloJulian DavidNessuna valutazione finora
- El Tronco EncefalicoDocumento17 pagineEl Tronco EncefalicoandytakerNessuna valutazione finora
- Exposicion Sistema Nervioso CentralDocumento28 pagineExposicion Sistema Nervioso CentralEmilio Jose Burbano RojasNessuna valutazione finora
- Embriogenesis Del Sistema NerviosoDocumento19 pagineEmbriogenesis Del Sistema NerviosoYoselin Quispe vilcaNessuna valutazione finora
- Neuroanatomia. Temario CompletoDocumento410 pagineNeuroanatomia. Temario CompletocetefloNessuna valutazione finora
- Sistema Nervioso CentralDocumento20 pagineSistema Nervioso CentralmelisadoraNessuna valutazione finora
- Wuolah-free-Tema 10. Tronco Del EncéfaloDocumento7 pagineWuolah-free-Tema 10. Tronco Del EncéfaloangiebenzoNessuna valutazione finora
- Resumen IIDocumento8 pagineResumen IICarla MálagaNessuna valutazione finora
- Anat. (17) Sistema Nervioso I 61 - 68Documento8 pagineAnat. (17) Sistema Nervioso I 61 - 68Andrea MontalvoNessuna valutazione finora
- Capitulo Nº11Documento41 pagineCapitulo Nº11Maribel CabreraNessuna valutazione finora
- El Sistema NerviosoDocumento7 pagineEl Sistema NerviosoLucia Fernandez AlonsoNessuna valutazione finora
- Apuntes Fundamentos BiologicosDocumento96 pagineApuntes Fundamentos BiologicosYu Jie VidondoNessuna valutazione finora
- Tallo CerebralDocumento26 pagineTallo CerebralAlbertoSanchezNessuna valutazione finora
- Breves Nociones de NeuroanatomiaDocumento14 pagineBreves Nociones de NeuroanatomiarocioruizlermosNessuna valutazione finora
- Apuntes Unificados Bases Biologicas Tercera UnidadDocumento23 pagineApuntes Unificados Bases Biologicas Tercera UnidadPao TarabataNessuna valutazione finora
- UNIFICADO BB2 3 y 4Documento39 pagineUNIFICADO BB2 3 y 4Pao TarabataNessuna valutazione finora
- Neuroanatomía Funcional (Universidad Nacional Mayor de San Marcos) Neuroanatomía Funcional (Universidad Nacional Mayor de San Marcos)Documento11 pagineNeuroanatomía Funcional (Universidad Nacional Mayor de San Marcos) Neuroanatomía Funcional (Universidad Nacional Mayor de San Marcos)Griselda LópezNessuna valutazione finora
- Resumen 1er Parcial NeuroDocumento19 pagineResumen 1er Parcial NeuroScarlet RiveraNessuna valutazione finora
- Tercer Parcial de NeuroDocumento10 pagineTercer Parcial de NeuroheliNessuna valutazione finora
- II Unidad Morfofisiologia III Completa Samuel ReyesDocumento252 pagineII Unidad Morfofisiologia III Completa Samuel ReyesDiego HernandezNessuna valutazione finora
- VT1 UF3 CompletaDocumento78 pagineVT1 UF3 CompletaJuan Ortiz GarciaNessuna valutazione finora
- PRÁCTICA 5 - ESCATE CHARCA JULISA CRISTELSISTEMA NERVIOSO - GrupoMiercoles - AYFA IIDocumento18 paginePRÁCTICA 5 - ESCATE CHARCA JULISA CRISTELSISTEMA NERVIOSO - GrupoMiercoles - AYFA IIJulisa Cristel Escate CharcaNessuna valutazione finora
- Sistema Nervioso Central: IntegrantesDocumento30 pagineSistema Nervioso Central: Integrantesrcandia806Nessuna valutazione finora
- Neuro Anatomia - Primer ParcialDocumento15 pagineNeuro Anatomia - Primer ParcialValeriaNessuna valutazione finora
- 2 SEMA TEO Organizacion Del Sistema NerviosoDocumento17 pagine2 SEMA TEO Organizacion Del Sistema NerviosoEsmeralda De la cruz fabianNessuna valutazione finora
- UT12. Sistema Nervioso y Órganos de Los SentidosDocumento24 pagineUT12. Sistema Nervioso y Órganos de Los SentidosNayaraNessuna valutazione finora
- Resumen NeuroanatomiaDocumento42 pagineResumen NeuroanatomiaAgustina BarrionuevoNessuna valutazione finora
- 1 Fisiología de Sistemas y CerebroDocumento32 pagine1 Fisiología de Sistemas y Cerebro《 V A E 》Nessuna valutazione finora
- Libro Fisiologí Del Sistema Nervioso-29-72Documento44 pagineLibro Fisiologí Del Sistema Nervioso-29-72Helen Nicol GarradoNessuna valutazione finora
- Sistema NerviosoDocumento23 pagineSistema NerviosoChristian GuerraNessuna valutazione finora
- NeuroanatomiaDocumento19 pagineNeuroanatomiaRodrigues Miranda Melina 8 8 6 0 8 1Nessuna valutazione finora
- Módulo de Neuroanatomía OKDocumento22 pagineMódulo de Neuroanatomía OKNacho MoralesNessuna valutazione finora
- 9 Tronco Encefalico Morfo (Autoguardado)Documento28 pagine9 Tronco Encefalico Morfo (Autoguardado)cristinaggonzNessuna valutazione finora
- Resumen Fina Parte 1Documento63 pagineResumen Fina Parte 1Maria Celina MassaNessuna valutazione finora
- Fisiología Tema 2Documento36 pagineFisiología Tema 2mts11violetNessuna valutazione finora
- Par CranealDocumento179 paginePar CranealRodrigo PonceNessuna valutazione finora
- TEMA 10. Encéfalo I. PROSENCÉFALODocumento14 pagineTEMA 10. Encéfalo I. PROSENCÉFALOrodrigomartinf97Nessuna valutazione finora
- 5 Base Del Cráneo y EncéfaloDocumento12 pagine5 Base Del Cráneo y EncéfaloRichard Álvarez ChungNessuna valutazione finora
- Anatomía Funcional Del Sistema Nervioso CentralDocumento5 pagineAnatomía Funcional Del Sistema Nervioso CentralRony Rodas CastellanosNessuna valutazione finora
- Tema 4Documento11 pagineTema 4NATALIANessuna valutazione finora
- Principales Subdivisiones Del Sistema NerviosoDocumento77 paginePrincipales Subdivisiones Del Sistema NerviosoBryanNessuna valutazione finora
- Sistema Nervioso PracticaDocumento14 pagineSistema Nervioso Practicaraul rojasNessuna valutazione finora
- NeuroembriologíaDocumento15 pagineNeuroembriologíaDanielaNessuna valutazione finora
- Práctica Disección EncéfaloDocumento6 paginePráctica Disección EncéfaloAlejandro Fernández MuñozNessuna valutazione finora
- Neuroanatomia Clinica de SnellDocumento7 pagineNeuroanatomia Clinica de SnellDavid BarzalloNessuna valutazione finora
- Neurocirugia 2Documento108 pagineNeurocirugia 2Jesus David Gonzalez MachinNessuna valutazione finora
- Spring DayDocumento3 pagineSpring DaysofiaNessuna valutazione finora
- Https 1Documento3 pagineHttps 1sofiaNessuna valutazione finora
- Teoría FractalDocumento3 pagineTeoría FractalsofiaNessuna valutazione finora
- Art. Las Psicoterapias de Benito Ezequiel - PsicoterapiaDocumento10 pagineArt. Las Psicoterapias de Benito Ezequiel - Psicoterapiakuniko alvaNessuna valutazione finora
- La Relacion Medico - Paciente PDFDocumento4 pagineLa Relacion Medico - Paciente PDFJohnM.GiraldoNessuna valutazione finora
- La Relacion Medico - Paciente PDFDocumento4 pagineLa Relacion Medico - Paciente PDFJohnM.GiraldoNessuna valutazione finora
- Aspectos Fisiologicos Acerca Del SueñoDocumento8 pagineAspectos Fisiologicos Acerca Del SueñoRifutokukuari Vilkiart RafikiNessuna valutazione finora
- La Lectura - 20170625225859Documento5 pagineLa Lectura - 20170625225859sofiaNessuna valutazione finora
- Aneurisma en El CerebroDocumento4 pagineAneurisma en El CerebrosofiaNessuna valutazione finora
- Bases Moleculares de La Regulación Ciclo CelularDocumento2 pagineBases Moleculares de La Regulación Ciclo CelularsofiaNessuna valutazione finora
- 1º Bachillerato. Física y QuímicaDocumento43 pagine1º Bachillerato. Física y QuímicaisabelNessuna valutazione finora
- Ats Trabajos en Espacios Confinados.Documento7 pagineAts Trabajos en Espacios Confinados.Ana Maria CalleNessuna valutazione finora
- Guia Del SenaDocumento10 pagineGuia Del SenaGloria Patricia Posada Perez100% (1)
- Guia Numeros y Algebra 1Documento6 pagineGuia Numeros y Algebra 1HernanMoragaNessuna valutazione finora
- EstratificacionDocumento9 pagineEstratificacionBrayan Pandal SaenzNessuna valutazione finora
- Metodo DeflexionDocumento8 pagineMetodo DeflexionJhonathan Romero PaucarNessuna valutazione finora
- Flujo en Medios PorososDocumento33 pagineFlujo en Medios PorososRené Fernando Gutiérrez CastilloNessuna valutazione finora
- Los Orígenes y Destinos de La Idea de Terceridad en El Psicoanálisis ContemporáneoDocumento31 pagineLos Orígenes y Destinos de La Idea de Terceridad en El Psicoanálisis ContemporáneoEnric BoleaNessuna valutazione finora
- Nivelacion9 Estadistica1PDocumento7 pagineNivelacion9 Estadistica1PPaula CortesNessuna valutazione finora
- Ventajas de La EncuestaDocumento3 pagineVentajas de La Encuestapromocion2010Nessuna valutazione finora
- Manual Pickit 3Documento19 pagineManual Pickit 3alvaroNessuna valutazione finora
- Taller Colaborativo 2Documento3 pagineTaller Colaborativo 2JuliAn NavarRoNessuna valutazione finora
- Cuatro Operaciones IDocumento2 pagineCuatro Operaciones IEnriqueMacoteCarhuas0% (1)
- WWW Hispasonic Com Tutoriales Sintesis 6 Fundamentos Sintesis Sustractiva I 38468Documento20 pagineWWW Hispasonic Com Tutoriales Sintesis 6 Fundamentos Sintesis Sustractiva I 38468DiegoArmandleBlancNessuna valutazione finora
- ADTD-32 Tratamiento de Datos Ausentes y AtípicosDocumento36 pagineADTD-32 Tratamiento de Datos Ausentes y Atípicosandreaaleman1818Nessuna valutazione finora
- MAGNETISMO Primera Edicion 2007Documento15 pagineMAGNETISMO Primera Edicion 2007nicolas dionisio ordonez barruetaNessuna valutazione finora
- RequisitosDocumento2 pagineRequisitosjeoberty peralta mendezNessuna valutazione finora
- Evaluacion Grado Once - SonidoDocumento1 paginaEvaluacion Grado Once - SonidoANDREA DEL PILAR CASTRILLON DIAZNessuna valutazione finora
- Segundo Examen Ciclo EspecialDocumento41 pagineSegundo Examen Ciclo EspecialJorge Cesar Rojas Ramos0% (1)
- Relación de Áreas para Quinto Grado de SecundariaDocumento4 pagineRelación de Áreas para Quinto Grado de Secundariajosecab1996Nessuna valutazione finora
- Introduccion A MatlabDocumento7 pagineIntroduccion A MatlabnelsonNessuna valutazione finora
- g.2 Vilaña - Mendez - Teresa Matematica IDocumento3 pagineg.2 Vilaña - Mendez - Teresa Matematica ITeresa MendezNessuna valutazione finora
- Grafica de La Función SenoDocumento11 pagineGrafica de La Función SenopriscilaNessuna valutazione finora
- 4 Guia de Trabajo Nº4Documento5 pagine4 Guia de Trabajo Nº4Catalina Fernanda Zambrano PaniaguaNessuna valutazione finora
- Probabilidad de Unión e Intersección de Sucesos 5°Documento9 pagineProbabilidad de Unión e Intersección de Sucesos 5°Dalmer Ignacio MattosNessuna valutazione finora
- Teorema Modificado de Castigliano1-SignedDocumento25 pagineTeorema Modificado de Castigliano1-SignedCuenta Netflixilbay2Nessuna valutazione finora
- AutoCAD 2D Referencias ExternasDocumento6 pagineAutoCAD 2D Referencias ExternasMaría BadilloNessuna valutazione finora
- Segmentos (Intermedio)Documento2 pagineSegmentos (Intermedio)Jhonny Davila PerezNessuna valutazione finora
- Ejm Coordenadas Geograficas A UtmDocumento9 pagineEjm Coordenadas Geograficas A UtmVictor Andres Sicus ChampiNessuna valutazione finora